КРОВЕНОСНАЯ СИСТЕМА
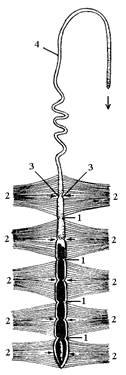
|
| Рис. 8. Спинной сосуд пчелы: 1 – клапаны сердца; 2 – крыловидные мышцы; 3 – щелевидные отверстия – остии; 4 – аорта (стрелками показано направление движения крови). |
Органы кровообращения. Кровеносная система насекомых незамкнутая. Она представлена спинным сосудом, который располагается в перикардиальном отделе тела и состоит из сердца и аорты (рис. 8). Сердце состоит из ряда камер. В каждой камере имеется по паре боковых отверстий – остий. Края отверстий завернуты, образуя остиальные клапаны. При диастоле они пропускают кровь в полость сердца, при систоле закрываются. Иногда края остальных клапанов вдаются в полость следующей камеры сердца и играют роль межкамерного клапана. Непосредственно под сердцем располагаются парные пучки мышц треугольной формы – крыловидные мышцы. Они входят в состав верхней диафрагмы и связаны с нижней стенкой сердца.
Пульсация сердца и диафрагм обеспечивает всасывание крови и ее продвижение по направлению к аорте. Из аорты кровь изливается в полость головы, а затем переходит в полость тела. Число сокращений камер сердца зависит от вида насекомого, его физиологического состояния, условий среды и колеблется от 15–30 до 150 сокращений в 1 мин.
Продвижению крови в различные придатки тела (усики, ноги, крылья, придатки брюшка) способствуют местные пульсирующие органы в виде ампул или сокращающихся перепонок, связанных с мышцами.
Кровь. Кровь насекомых, или гемолимфа, состоит из жидкой плазмы и клеточных элементов, или кровяных телец, гемоцитов.Плазма обычно бесцветна или окрашена в зеленоватый цвет. Лишь у живущих в илистом грунте личинок комаров звонцов, или мотылей, плазма окрашена в красный цвет в связи с присутствием вещества, по биохимическому составу достаточно близкого к гемоглобину крови позвоночных.
Плазма содержит неорганические соли, белки, аминокислоты, углеводы, жиры, мочевую кислоту, ферменты, гормоны, пигменты, воду 75–90%, рН гемолимфы 6–7. Гемоциты имеют мезодермальное происхождение. Форма различна – подвижные и неподвижные гемоциты. В 1 мм3 гемолимфы божьей коровки содержится до 80 тыс. гемоцитов. Различают 3 типа гемоцитов: 1 – хромофильные лейкоциты; 2 – амебоциты и гемоциты с зернистой и гомогенной плазмой; 3 – фагоциты.
Функции гемолимфы: 1) разнос по телу питательных веществ и снабжение ими тканей; 2) поглощение из тканей вредных продуктов обмена и перенос их к органам выделения; 3) так как гемолимфа содержит гормоны, выделяемые железами внутренней секреции, то она регулирует физиологические процессы; 4) механическая: создает необходимое внутреннее давление, или тургор; 5) защитная функция: выбрызгивается кровь для самозащиты (кузнечики, саранча) или содержит сильно действующие биологически активные вещества и выделяют их наружу при опасности.
В то же время дыхательная функция крови незначительна, так как из-за отсутствия гемоглобина емкость гемолимфы невелика и ограничена главным образом количеством растворенного в ней кислорода.
ОРГАНЫ ДЫХАНИЯ
Дыхательная система состоит из дыхалец (стигм), трахей и трахеол (рис. 9). Дыхание насекомых осуществляется через систему трахей, распространенных по всему телу, реже – через поверхность кожных покровов. Трахеи представлены полыми трубками, выстланными хитином в виде спиральных утолщений, препятствующих спаданию трахей при движении и изгибах тела. Трахеи разветвляются на мельчайшие капилляры – трахеолы диаметром менее 1 мкм, доставляющие кислород воздуха непосредственно к тканям и клеткам тела.
Снаружи трахеи открываются парными дыхальцами, расположенными по бокам тела. У большинства взрослых насекомых и у личинок насекомых с неполным превращением развито десять пар дыхалец – две пары на груди и восемь пар на брюшке. Но у ряда групп высших насекомых и особенно их личинок и куколок число дыхалец сокращается или они отсутствуют (первичнобескрылые, личинки паразитических и водных насекомых). Дыхальца устроены сложно: отверстие, затем кольцеобразный склерит (рама), затем особая камера – атриум, замыкающий аппарат, который может быть наружным или внутренним. Атриум лишен спиральной выкладки, служит входом в стигмальную трахею и снабжен фильтрующим аппаратом, предохраняющим дыхательную систему от засорения (рис. 10). У прямокрылых, жуков, пчелиных, двукрылых и некоторых других летающих насекомых развиты воздушные мешки, представляющие местное расширение трахейных стволов, идущих от дыхалец. Они лишены спиральных утолщений, и полость их может спадаться при выдохе воздуха.
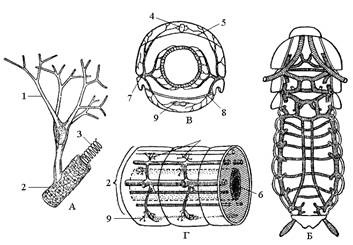
Рис. 9. Трахейная система. А – окончание трахеи с трахеолами; Б – часть главнейших стволов трахей у черного таракана; В – схема поперечного ветвления трахей в сегменте; Г – схема продольных стволов трахей в сегментах: 1 – трахеолы, 2 – трахеи, 3 – тенидии, 4 – сердце, 5 – верхняя диафрагма, 6 – кишечник, 7 – стигма, 8 – нижняя диафрагма, 9 – нервная цепочка.
Дыхание. Поступление воздуха в трахейную систему происходит чаще всего активно, с помощью дыхательных движений. При этом те или иные дыхальца открываются или закрываются, выполняя вдох или выдох. Ритм дыхательных движений зависит от вида насекомого, его состояния и внешних условий. Так, медоносная пчела в покое совершает около 40 дыхательных движений в 1 мин, а в движении – до 120; у некоторых саранчовых увеличение их числа от 6 до 26 и более происходит при повышении температуры среды от 00 до 27 0С и выше.
У многих видов насекомых воздух вдыхается через грудные и выдыхается через брюшные дыхальца. Ритм работы дыхалец связан с дыхательными движениями брюшка; при повышении или понижении давления воздуха, вызываемых этими движениями, одни дыхальца
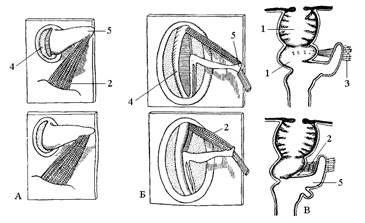
Рис. 10. Дыхальца и их замыкательный аппарат. А – брюшное дыхальце клопа; Б – дыхальце гусеницы бабочки; В – брюшное дыхальце муравья, сверху открытое, снизу закрытое: 1 – атриум; 2 – закрывающая мышца; 3 – открывающая мышца; 4 – рама; 5 – замыкающий рычаг.
открываются наружу, другие – внутрь тела насекомого. Однако под влиянием больших доз углекислого газа, различных ядов, а иногда и без видимых причин циркуляция воздуха может измениться, т.е. он начинает поступать через брюшные дыхальца и выходить через грудные. Кроме этого, при повышении содержания углекислого газа и недостатке кислорода в окружающей среде дыхальца остаются открытыми более продолжительное время, в связи с чем фумигация помещения против вредителей будет более эффективной.
Дыхание представляет собой окислительный процесс, протекающий за счет потребления кислорода и выделения углекислоты. Процесс окисления идет при участии окислительных ферментов – оксидаз и сопровождается постепенным расщеплением молекул расходуемых соединений – углеводов, жиров, белков – и выделением энергии. Расщепление этих соединений в конечном счете завершается образованием углекислого газа и воды, а для белков еще и появлением продуктов распада, связываемых в более безопасные для организма соединения типа мочевины и ее солей. Освобождающаяся при этом тепловая и механическая энергия идет на поддержание жизнедеятельности организма. Этим объясняется физиологическая необходимость дыхания.
Таким образом, дыхание сопровождается газообменом. Процесс газообмена характеризуется дыхательным коэффициентом(ДК), представляющим отношение выделенного углекислого газа к общему количеству поглощенного кислорода. По этому показателю можно судить, какие вещества используются в данный момент в качестве источника энергии. При окислении углеводов ДК = 1, так как количество поглощенных молекул кислорода и выделенных молекул углекислого газа равно между собой: С6Н12О6 + 6О2 = 6CО2 + 6Н2О, а по закону Авогадро объемы этих газов равны. При использовании менее окисленных соединений жиров ДК снижается до 0,7, а белков – до 0,77–0,82. Например, при голодании тараканов ДК снижается до 0,65–0,85, что соответствует преимущественному расходованию ранее запасенных жиров.
Температура тела. Насекомые относятся к животным с непостоянной температурой тела. Она зависит от интенсивности процессов образования тепла и его отдачи. Источниками образования тепла у насекомых являются, с одной стороны, процессы обмена веществ в организме, сопровождающиеся выделением тепловой энергии, и лучистая энергия солнца или нагретый им воздух – с другой.
По данным И.Д. Стрельникова, температура тела насекомых, находящихся в покое и не подвергающихся облучению солнцем, примерно равна температуре окружающей среды. В связи с тем, что температурный оптимум для многих видов колеблется около 20–35 0С, насекомые могут в известных пределах регулировать температуру тела за счет изменения мышечной активности (движение, полет) или передвижения на более нагретые или прохладные участки, иногда за счет изменения позы. Весомое значение в регуляции температуры тела могут иметь испарение воды с поверхности кожных покровов и вентиляция трахей, особенно с помощью воздушных мешков.
ОРГАНЫ ВЫДЕЛЕНИЯ
Выделение различных веществ организмом насекомого складывается из трех процессов – экскреции, секреции и инкреции, или внутренней секреции. В соответствии с этим различают экскреторные органы, экзокринные и эндокринные железы. Общее свойство – способность выделять наружу или внутрь разнообразные вещества и тем самым участвовать в обмене веществ в организме. В одном случае образуются вредные или бесполезные продукты обмена веществ и они удаляются из организма, а в другом случае разнообразные органы и железы выделяют необходимые для нормальной жизнедеятельности вещества.
Экскреторные органы и экскреция. Экскреция – процесс выделения ненужных и вредных для организма веществ – экскретов, образующихся в процессе обмена веществ вне пищеварительной системы. К ним относятся растворимые продукты азотного обмена в виде мочевой кислоты и ее солей, щавелевая и фосфорная кислоты, катионы калия, натрия и пр. Экскреторная или выделительная система поддерживает относительное биохимическое постоянство внутренней среды и контролирует ионный состав крови. Вредные вещества, удаляемые из организма, называются экскретами, а сам процесс – экскрецией. Экскреторные органы делят на две группы: выводящие экскреты и накапливающие их в клетках. К первой группе относятся мальпигиевы сосуды и нижнегубные железы некоторых насекомых, ко второй – уратные клетки жирового тела.
Мальпигиевы сосуды – основной экскреторный орган. Они представляют слепые на свободном конце трубочки, впадающие в кишечник впереди или позади пилорического клапана, т.е. на границе между средним и задним отделом кишечника Стенки мальпигиевых сосудов состоят изнутри из одного слоя эпителиальных клеток, снаружи одеты базальной перепонкой и иногда имеют собственную мускулатуру, обеспечивающую им подвижность (см. рис. 2).Количество трубочек колеблется от двух (червецы) до 250 (пустынная саранча). Однако, несмотря на резкие различия в числе трубочек, величина их поверхности, приходящаяся на единицу массы, довольно близка. Так, 60 трубочек таракана имеют поверхность 132000 мм2, что составляет 412 мм2 на 1 мг массы тела, тогда как поверхность шести сосудов бабочки из семейства коконопряды составляет 209000 мм2, или 500 мм2 на 1 мг массы тела.
В процессе контакта гемолимфы со стенками мальпигиевых сосудов водные растворы мочекислых солей калия и натрия (ураты) проникают в полость концевого отдела сосудов. По пути в основной отдел имеющаяся в мальпигиевом сосуде свободная углекислота вытесняет мочевую кислоту из уратов, в результате чего образуются карбонат калия и свободная мочевая кислота. Последняя по мере повышения ее концентрации выпадает в виде мелких кристаллов и выводится из сосуда в кишечник, а затем через анальное отверстие вместе с экскрементами задней кишки – наружу. Легкорастворимый карбонат калия вместе с водой поступает через стенки сосуда снова в кровь, встречает там мочевую кислоту, образует ураты, которые снова попадают в полость мальпигиева сосуда, и т.д.
Кроме экскреторных мальпигиевы сосуды могут выполнять и дополнительные, секреторные, функции. У личинок сетчатокрылых и некоторых жуков они выделяют секрет, используемый для образования кокона или для цементирования кокона солями кальция. У жужелиц и некоторых других жуков мальпигиевы сосуды выделяют также пищеварительный фермент дипептидазу.
Экскреторными функциями обладают нижнегубные, или лабиальные, железы у некоторых первичнобескрылых (ногохвосток, щетинохвосток). Они расположены в голове в виде парных образований и имеют общий канал, открывающийся у основания нижней губы.
К экскреторным органам, накапливающим экскреты, относятся уратные клетки жирового тела. Экскреты откладываются в виде кристаллов мочевой кислоты, свободно взвешенных в цитоплазме. Отложение мочевой кислоты в жировом теле может происходить наряду с обычной ее экскрецией через мальпигиевы сосуды. Однако у личинок и куколок внутренних паразитов насекомых вынос экскретов наружу происходит лишь после вылета имаго. Выведение их в организм хозяина в более ранний период могло бы привести к отравлению и гибели самого паразита.
Экзокринные железы и секреция. Секреция – процесс выделения различных веществ, или секретов, нужных организму. К экзокринным железам, осуществляющим этот процесс, относятся уже отмеченные выше: 1) слюнные железы и железистый эпителий средней кишки, выделяющие пищеварительные ферменты, восковые, лаковые, шелкоотделительные железы, секрет которых служит для механической защиты тела, и 2) железы, выделяющие биологически активные вещества, являющиеся средством химического воздействия на других животных. Последние Я.Д. Киршенблат назвал телергонами.Железы, выделяющие телергоны, имеют протоки, выходящие на поверхность тела или в полости, связанные с внешней средой. Телергоны подразделяются на гетеротелергоны, воздействующие на другие виды животных, и телергоны, или феромоны, воздействующие на особей своего вида.
К гетеротелергонам относятся вещества химической защиты насекомого от врагов (яд жалящих перепончатокрылых – пчел, ос), а также сильнопахнущие и другие отпугивающие вещества – репелленты (секрет желез жука-бомбардира, взрывающийся на воздухе).
К феромонам относятся вещества, определяющие концентрацию насекомых при использовании источников пищи или при поиске мест для спаривания, – феромоны скучивания, выявленные у прямокрылых, жуков-короедов; феромоны тревоги, или обороны, – у термитов, тлей, жалящих перепончатокрылых; феромоны, контролирующие развитие половых желез у рабочих пчел; половые феромоны, или половые аттрактанты, привлекающие особей другого пола.
Половые феромоны выделяются в ничтожно малых количествах и воспринимаются другим полом на значительных расстояниях. Так, секрета одной неоплодотворенной самки часто бывает достаточно для привлечения десятков особей самцов того же вида, расположенных от нее на расстоянии десятков, а иногда сотен метров. Некоторые половые феромоны уже синтезированы и используются в борьбе с отдельными видами вредных насекомых или для учета их появления и численности на полях и в садах.
Эндокринные железы и инкреция. Инкреция, или внутренняя секреция, представляет собой процесс выделения секретов эндокринных желез, или гормонов, непосредственно в кровь. Они транспортируются кровью по всему телу и регулируют процессы обмена веществ и развития насекомых. В целом эндокринные железы регулируют обменные процессы и развитие насекомых и связанные с ними явления – личиночный рост, линьки, торможение, развитие (диапауза), половое созревание, поведение, изменение окраски тела. Наиболее хорошо изучены четыре группы эндокринных желез у насекомых: нейросекреторные клетки головного мозга,кардиальные тела (рис. 11), переднегрудные, или проторакальные, железы и прилежащие телá.
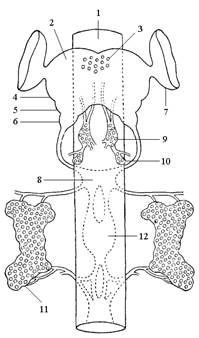
|
| Рис. 11. Схема эндокринной системы насекомых и переднего отдела центральной нервной системы: 1 – кишечник; 2 – надглоточный узел; 3 – нейросекреторные клетки головного мозга; 4 – протоцеребрум; 5 – дейтоцеребрум; 6 – тритоцеребрум; 7 – глазные доли; 8 – подглоточный узел; 9 – кардиальные тела; 10 – прилежащие тела; 11 – переднегрудная железа; 12 – ганглии брюшной нервной цепочки. |
Нейросекреторные клетки головного мозгаи других ганглиев выделяют группу физиологически активных веществ – нейрогормонов, участвующих в регуляции развития, поведения и обмена веществ. Нейросекрет синтезируется в клетках, а затем транспортируется по аксонам в прилежащие и кардиальные тела, накапливается здесь и выделяется в гемолимфу. Нейросекреторные клетки образуют единую нейросекреторную систему совместно с кардиальными телами – играющую роль в гормональной системе насекомых. Мозговой гормон регулирует деятельность всех эндокринных органов, обладает активационной функцией, играет роль пускового механизма, воздействующего на прилежащие и кардиальные тела и переднегрудные железы. К несколько более изученным нейрогормонам от-носятся: эмбриональной диапаузы, диуретический, гипергликемический, адипокинетический, активационный.
Так, гормон эмбриональной диапаузы, выделенный в гемолимфу куколок и бабочек тутового шелкопряда, переходит в яйцеклетку и вызывает эмбриональную диапаузу. Диуретический гормон усиливает экскрецию первичной мочи мальпигиевыми сосудами, гипергликемический – повышает содержание в гемолимфе дисахарида трегалозы, накапливающейся в жировом теле, адипокинетический – стимулирует распад триглицеридов в жировом теле, освобождая диглицеридную транспортную форму жиров, поступающих из трофоцитов в гемолимфу.
Из нейрогормонов в большей степени изучен активационный гормон. Он образуется в нейросекреторных клетках переднего отдела головного мозга – протоцеребрума (рис. 12) – и активизирует деятельность другой группы эндокринных желез – проторакальных, выделяющих линочный гормон, или экдизон. Личинка клопа Rhodnius prolixus линяет через 12–28 дней после питания кровью. Если ее обезглавить не позднее критического периода, т.е. за 4 дня для младших и за 7 дней для старшего (V) возраста, то линька не произойдет, хотя в таком состоянии насекомое может жить в течение года. Следовательно, активационный гормон через посредство другого гормона – экдизона обеспечивает процессы линьки и метаморфоза.
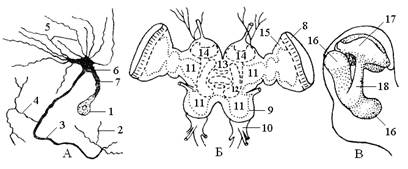
Рис. 12. Элементы нервной системы насекомых: а – нейрон; б – головной мозг саранчи; в – схема грибовидного тела мозга таракана; 1 – нервная клетка; 2 – концевые разветвления аксона; 3 – аксон; 4 – его коллатеральная ветвь; 5 – дендриты; 6 – главное мозговое разветвление; 7 – клеточный отросток; 8 – протоцеребрум; 9 – дейтоцеребрум; 10 – тритоцеребрум; 11 – нейропиль; 12 – центральное тело; 13 – протоцеребральный мост; 14 – грибовидные тела; 15 – зрительная доля; 16 – лопасти ножки грибовидного тела; 17 – чашечка грибовидного тела; 18 – ножка грибовидного тела.
Кардиальные тела расположены впереди прилежащих тел, связаны нервами с головным мозгом и свойственны личинкам имаго. Они регулируют дыхательный обмен у тараканов, стимулируют образование липидов в жировом теле у насекомых саранчовых.
Проторакальные железы расположены по бокам переднегрудного ганглия, или узла, брюшной нервной цепочки. Железы выделяют линочный гормон, или экдизон, который стимулирует линьку и метаморфоз у личинок и прекращает состояние диапаузы. Установлено, что проторакальные железы неактивны в межлиночный период, умеренно активны в период линек и достигают максимальной активности перед окукливанием. Экдизон неспецифичен, т.е. может действовать на различные виды насекомых.
Прилежащие тела (corpora allata) расположены над пищеводом позади головного мозга в виде пары округлых желез. Они выделяют ювенильный гормон, или неотении. Гормон препятствует превращению личинки во взрослое насекомое, т.е. является ингибитором метаморфоза. Он выделяется в кровь периодически, в связи с чем изменяется соотношение между ювенильным гормоном и экдизоном и происходят линьки. К концу периода личиночного развития содержание ювенильного гормона в крови резко уменьшается и наступает метаморфоз. Создание высокой концентрации гормона в организме личинки в этот период, например при искусственных инъекциях, может вызвать дополнительные линьки и чрезмерный рост личинок.
К другим функциям ювенильного гормона относят стимулирование процессов репродуктивного развития, особенно образование желтка в половых железах самок и развитие придаточных желез самца после метаморфоза, появление кастового полиморфизма у общественных насекомых, фазового полиморфизма у саранчовых, сезонного полиморфизма у тлей. За последние годы не только изучена химическая природа экдизона и ювенильного гормона, но и синтезированы многочисленные аналоги последнего, названные ювеноидами. Ряд ювеноидов испытывают в борьбе с различными насекомыми. Изучают также антиювенильные гормоны растительного происхождения – прекоцены, вызывающие преждевременный метаморфоз личинок некоторых видов клопов во взрослых насекомых, неспособных к воспроизводству потомства.
ОРГАНЫ РАЗМНОЖЕНИЯ
Половая система выполняет функцию размножения и тем самым обеспечивает существование вида, т.е. это система органов видовой жизни. Как правило, насекомые раздельнополы.
Половая система самки. Половая система самки включает парные яичники, парные яйцеводы, непарный яйцевод, парные придаточные железы, семяприемник, иногда яйцеклад (рис. 13).
Яичники состоят из яйцевых трубок, в которых формируются яйца. Число яйцевых трубок у разных видов насекомых варьируется от четырех-восьми пар у некоторых жуков и бабочек до 220 пар у медоносной пчелы и до 12000 пар у самки термита.
Яйцевая трубка подразделена на вершинную часть – гермарий и основную – вителлярий. В гермарий из первичных половых клеток оогоний образуются ооциты и питательные клетки. Созревшие ооциты превращаются в яйца и поступают в вителлярий. Здесь по мере созревания они увеличиваются в размере, часто отделяясь друг от друга сужением или перехватом; в этих случаях вителлярий оказывается разделенным на ряд последовательно утолщающихся яйцевых камер. По окончании развития питание яйца прекращается, и фолликулярный эпителий выделяет хитинообразную оболочку яйца – хорион.
В зависимости от характера поступления питательных веществ в формирующиеся яйца яйцевые трубки делят на три типа: паноистический, телотрофический, политрофический. В трубках паноистического
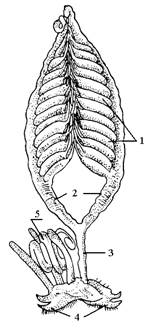
|
| Рис. 13. Половая система самки перелетной саранчи: 1 – яичник; 2 – яйцеводы; 3 – непарный яйцевод; 4 – яйцеклад; 5 – семяприемник. |
типа дифференцированные питательные клетки отсутствуют, питание осуществляется фолликулярным эпителием (стрекозы, представители отрядов надотряда ортоптероидных). В трубках телотрофического типа питательные клетки остаются в гермарии и образуют многоядерную трофическую камеру. Пищевые продукты транспортируются из камеры по длинным цитоплазматическим тяжам, проникающим непосредственно в ооциты (равнокрылые, клопы и жуки из подотряда разноядных). Трубки политрофического типа отличаются наличием обособленных питательных клеток, чередующихся с яйцевыми клетками (плотоядные жесткокрылые, сетчатокрылые, чешуекрылые и др.).
Созревшие яйца поступают в парные яйцеводы, затем в непарный яйцевод, куда впадает проток семяприемника. В это время сперматозоиды выходят из семяприемника и проникают в яйцо. Иногда у самок непарный семяпровод расширяется на заднем конце, образуя совокупительную сумку – влагалище. При копуляции сперматозоиды попадают сюда, а затем в семяприемник. У большинства чешуекрылых совокупительная сумка имеет независимое копулятивное отверстие, и тогда второе отверстие, из непарного яйцевода, служит лишь для вывода яиц наружу.
В непарный яйцевод открывается и проток придаточных желез, выполняющих разнообразные функции. Они выделяют секрет для приклеивания яиц к субстрату у бабочек, служат для образования яйцевого кокона – оотеки у тараканов и богомолов или кубышки у саранчовых и т.д.
Половая система самца. Органы размножения самца состоят из парных семенников, парных семяпроводов, непарного семяизвергательного канала, придаточных половых желез и копулятивного органа, или фаллуса. В состав семенника входит от одного до нескольких десятков семенных трубочек, или фолликулов, где размножаются зародышевые клетки и формируются сперматозоиды.
Созревшие сперматозоиды из семенников поступают в семяпроводы, концы которых нередко расширяются, образуя семенные пузырьки. Отсюда они поступают в семяизвергательный канал, который выталкивает сперму через копулятивный орган при спаривании.
Придаточные железы в числе от одной до трех пар впадают в семяизвергательный канал. Их секрет защищает сперму от внешних воздействий при спаривании (например, у пчелиных). У многих прямокрылых, богомолов, некоторых жуков секрет придаточных желез обволакивает порцию спермы, образуя своеобразную капсулу, называемую сперматофором. При спаривании самец вводит сперматофор в половое отверстие самки или прикрепляет его к нему, затем сперматозоиды переходят из сперматофора в половые пути самки.
Оплодотворение – важнейшая сторона биологии размножения. У насекомых оплодотворение может быть различным. Живущие во влажной среде спаривания самцы рассеивают капельки спермы или сперматофоры в местах обитания вида. Самка, встречая на пути порции спермы, захватывает ее половым отверстием (низшие насекомые). У высших насекомых сперматофора вводится в половые пути самки. Затем самка приступает к откладке яиц (может быть живорождение).
ЛИТЕРАТУРА
1. Осмоловский, Г.Е. Энтомология / Г.Е. Осмоловский, Н.В. Бондаренко. Л.: Колос, 1980.
2. Бей-Биенко, Г.Я. Общая энтомология / Г.Я. Бей-Биенко. М.: Высш. шк., 1971.
3. Вредители сельскохозяйственных культур и лесных насаждений / под ред. В.П. Васильева. Киев, 1973. Т. 1; 1974. Т. 2; 1975. Т. 3.
Дата добавления: 2017-11-21; просмотров: 1108;
Поиск по сайту
Узнать еще
- Arthropoda. Клещи. Систематика. Морфология. Медицинское значение.
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- D) Система класифікації за підтримкою багато гілкового виконання програми.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
