Структура генов эукариот
Позже, когда исследователи начали изучать гены различных белков в клетках эукариот, обнаружилось, что взаимодействие генов и белков в этих организмах более сложное, чем взаимодействие генов и белков прокариот. Первые примеры такого взаимодействия были получены в 1977 году в лабораториях Филиппа Шарпа и Пьера Шамбона. Вместе со своими коллегами они гибридизировали мРНК различных генов с теми ДНК, с которых были сняты эти информационные копии. У бактерий последовательность мРНК идентична последовательности кодирующей цепи ДНК (за исключением того, что место тимина занимает урацил), поэтому структура гибридных молекул была достаточно проста.
Но когда под электронным микроскопом были сделаны снимки гибридных молекул генов эукариот, то в них обнаружился ряд петель. Это значит, что мРНК и ДНК имеют не совсем идентичную последовательность, и петли были как раз теми местами, в которых они не могли соединяться. Когда последовательность мРНК сравнили с последовательностью ДНК, стало понятно, что кодирующая последовательность генов в некоторых местах прерывается некодирующей последовательностью, то есть некоторые нуклеотиды не кодируют синтез белка. Впоследствии выяснилось, что это типичная картина для ДНК эукариот.
Кодирующая последовательность гена называется экзоном, а некодирующая последовательность — интроном. Некоторые гены имеют в своей структуре несколько интронов. Часто обнаруживают и такие гены, в которых больше интронов, чем экзонов.
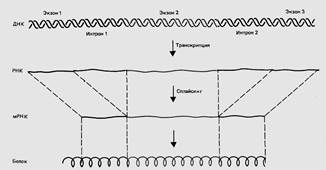
Рис. 2. Прерывистые гены экспрессируются в виде РНК- предшественника, из которой затем удаляются интроны; экзоны, соединяясь вместе, образуют мРНК.
В общем случае при транскрипции генов эукариот образуются большие молекулы РНК, содержащие как экзоны, так и интроны. Таким образом, полученные транскрипты, являются гетерогенными, т.е. они не на всем протяжении способны к трансляции. Поэтому первичные транскриптоны эукариот называются гетерогенной ядерной РНК (гяРНК) или про-мРНК.
В дальнейшем происходит преобразование про-мРНК, или ее «созревание», при котором она еще в ядре вовлекается в процессинг,состоящий в том, чтоособые комплексы ферментов (сплайсингсомы) вырезают из транскрипта все интроны. Затем экзоны соединяют в одну мРНК, кодирующую производство белка. Процесс соединения экзонов называется сплайсингом (сращивание).
В результате процессинга образуется «зрелая» мРНК, которая по размерам значительно короче, чем про-РНК. Молекулы про-мРНК содержат, в среднем, более 50 000 нуклеотидов, а созревшая мРНК – 500-3000 нуклеотидов. Далее полученная м-РНК транслируется как обычно. Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот.
Причины, по которым природа придерживается такой структуры, до сих пор не ясны, но ее можно объяснить как с эволюционной точки зрения, так и с точки зрения развития организма. Если говорить об эволюции, то такая структура ценна тем, что позволяет экспериментировать с генами и создавать новые гены. Кроссинговер может происходить внутри интронов, и в таком случае ошибки будут несущественными, а при рекомбинации могут образоваться новые экзоны и как следствие новые белки. Часто бывает так, что отдельный экзон кодирует отдельную область, или домен, белка, то есть отдельную часть белка с особыми функциями. Поэтому включение в ген нового экзона приведет к созданию белка с новыми областями и, возможно, с новыми функциями. Такое изменение генетической структуры может служить источником эволюции.
Другой особенностью работы генов эукариот является то, что сплайсинг мРНК, полученной с одного и того же локуса ДНК, может происходить по-разному. Это явление получило название альтернативный сплайсинг и оно приводит к тому, что конечный продукт –«созревшая» мРНК имеет разную нуклеотидную последовательность.
Сущность альтернативного сплайсинга заключается в том, что одни и те же участки исходной мРНК могут быть и интронами, и экзонами. Тогда одному и тому же участку ДНК соответствует несколько типов зрелой мРНК и, соответственно, несколько разных форм одного и того же белка.
На рис.3 представлен вариант альтернативного сплайсинга участков гена эукариот. В первом случае экзон 1 и экзон 2 соединяются вместе, а участки, обозначенные «сегмент» и «интрон», удаляются из РНК. Во втором случае соединяются экзон 1, сегмент и экзон 2, а удаляется только интрон. Таким образом, область, обозначенная «сегмент», служит интроном в первом случае и экзоном - во втором. В результате этих двух типов сплайсинга образуются два белка, которые имеют одинаковые концы, но различаются по средней части. Так одна последовательность ДНК может кодировать более одного белка.
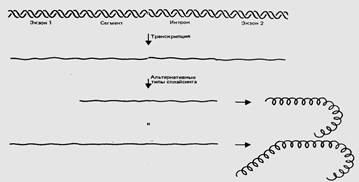
Рис 3. Гены, имеющие альтернативные пути экспрессии, бывает трудно разграничить.
Существует также транс–сплайсинг, который заключается в соединение экзонов, кодируемых разными генами (иногда даже из разных хромосом), в одну зрелую молекулу мРНК.
Таким образом, структура интрон-экзон позволяет одной нуклеотидной последовательности кодировать синтез более одного белка.
Гены эукариот, в силу их более сложной организации и функционирования, в результате чего получаются разные продукты-полипептиды, получили название «транскриптоны».
Структура гена в полной мере проявляется в свойствах белков. Это конечное проявление структуры генов -синтез полипептидов по созданной мРНК, называется проявлением генов.
Дата добавления: 2017-05-02; просмотров: 4700;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- Cоединения галогенов
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- II.1.2. Гены. Структура антигенов
- MS Word: проверка орфографии, поиск и замена, режимы просмотра документа (обычный, разметки страницы, структура).
- VIII. Формирование и структура характера
- А) пространственная структура.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
