Редуценты и детритофаги
ТРОФИЧЕСКАЯ СТРУКТУРА БИОЦЕНОЗА. ПИЩЕВЫЕ ЦЕПИ
Главная функция биоценозов в природе – поддержание круговорота вещества. Этот процесс базируется на пищевых взаимоотношениях видов.
Именно на этой основе органические вещества, синтезированные автотрофными организмами, претерпевают многократные химические трансформации и в конечном итоге возвращаются в среду в виде неорганических продуктов, вновь вовлекаемых в круговорот. Именно поэтому, при всем многообразии видов, входящих в состав различных экологических сообществ, каждое сообщество непременно включает представителей всех трех принципиальных экологических групп организмов:
- продуцентов,
- консументов и
- редуцентов.
В конкретных сообществах продуценты, консументы и редуценты представлены популяциями многих видов, состав которых специфичен для каждого отдельного сообщества. Однако все виды распределяются по нескольким группам в зависимости от места в общей системе круговорота вещества и потоков энергии. Равнозначные в этом плане виды образуют определенные трофические уровни, а взаимоотношения между видами разных уровней называют системой цепей питания. Совокупность трофических цепей в их конкретном выражении, в том числе прямые и косвенные взаимоотношения видов, формируют целостную трофическую структуру сообщества или биоценоза.
Трофические уровни
Уровень первичной продукции образует группа видов-продуцентов. На этом уровне утилизируется внешняя энергия и создается масса органического вещества.
Первичные продуценты – это основа трофической структуры всего существования сообщества.
Таксономический состав уровня. Продуценты представлены растениями (за исключением редких бесхлорофилльных форм), а также фотоавтотрофными прокариотами (водорослями). В особых случаях (например, в глубоководных системах) в качестве первичных продуцентов выступают бактерии хемосинтетики.
Биомасса органического вещества, синтезированного автотрофами, является первичной продукцией экосистемы. Скорость, с которой формируется эта биомасса, называется биологической продуктивностью экосистемы.
Продуктивность выражается количеством биомассы (или ее энергетическим эквивалентом), синтезируемой за единицу времени. Общая сумма биомассы рассматривается как валовая продукция, а та ее часть, которая возникла в результате прироста – как чистая продукция. Разница между валовой и чистой продукцией определяется затратами энергии на жизнедеятельность, т.е. на собственное поддержание экосистемы. Например, в умеренном климате эти различия могут составлять 40-70% валовой продукции.
Чистая первичная продукция, накопленная в виде биомассы организмов-атотрофов, служит источником питания для представителей следующих трофических уровней.
Консументы 1го порядка. Это непосредственные потребители первичной продукции.
Состав. В наиболее типичных случаях, когда первичная биомасса продуцирована автотрофными организмами, консументы 1го порядка представлены растительноядными животными – фитофагами. Таксономический и экологический состав фитофагов весьма разнообразен и приспособлен к питанию разными видами растительного корма.
Среди фитофагов различают семеноядных (зерноядных) животных, листоядных, травоядных. Выделяются животные, питающиеся преимущественно почками деревьев и кустарников, их побегами. Обычно отдельно указывается на группы животных-фитофагов, питающихся лишайниками (лемминги) и ветошью (т.е. засохшими частями в основном травянистых растений – оленьи). Есть и такие фитофаги, как нектарники – колибри, пчелы. Выделяют и специальную группу ксилофагов – питающихся преимущественно древесиной (жуки, термиты, частично слоны и т.д.).
Фитофаги обладают следующими морфологическими приспособлениями к потреблению пищи:
- у них эволюционно сформировался грызущий тип ротового аппарата и
- различного рода приспособления к измельчению, перетиранию пищи.
Такая морфология – результат адаптации к особенностям пищи: во-первых, растения обычно прикреплены к субстрату и, во-вторых, растения обладают прочными тканями.
Зубные системы грызущего или перетирающего типов имеют многие растительноядные млекопитающие. Грызущий ротовой аппарат свойственен и насекомым.
У растительноядных карповых рыб над глоточными зубами, на нижней конечности мозгового черепа имеется особое образование – «жерновок», который представлен ороговевшей поверхностью и предназначен для механического измельчения пищи.
Некоторые животные приспособлены к питанию соком растений или нектаром цветов (пчелы, колибри). У них ротовой аппарат устроен в виде трубочки, с помощью которой высасывается жидкая пища. Эта пища богата высококалорийными, легкорастворимыми веществами.
У птиц-фитофагов для измельчения заглоченной растительной пищи развился мускульный желудок, особенно хорошо выраженный у зерноядных видов. Такое приспособление вынуждает растительноядных птиц постоянно заботиться о гастролитах. Гастролиты – песок или мелкие камешки, которые птицы собирают, заглатывают и аккумулируют в мускульных желудках. Эти камешки являются «жерновами», с помощью которых размалывается пришедшая в желудок пища. Гастролиты не вечны, от этой работы они стираются и, кроме того, растворяются желудочными соками. К тому же птицы, похоже, способны при необходимости избавляться от гастролитов, выводя их в кишечник и далее наружу (через клоаку). Очевидно, что у них существует потребность систематически пополнять запас гастролитов.
Решать проблему пополнения желудка гастролитами порой бывает непросто, особенно в равнинных районах, где грунты обычно покрыты мощными травяными или моховыми «подушками». Дополнительные проблемы создает снежный покров. В результате таких ограничений у целой группы птиц (например, у курообразных) сформировались сезонные и региональные особенности поведения, связанные с «галькованием» (т.е. со сбором гальки). В основном эти особенности касаются размещения птиц. В частности в период массового сбора гастролитов (например, осенью, перед самыми снегопадами) они концентрируются неподалеку от галечников, т.е. от открытых грунтов, богатых галькой. В равнинных районах галечники обычно располагаются в поймах рек и по разнообразным карьерам. Нередко в качестве галечников птицами используются проселочные дороги.
Приспособления к питанию растениями обнаруживаются и на физиологическом уровне. Наиболее ярко такие приспособления выражены у животных, питающихся НЕрепродуктивными органами растений, а грубыми тканями вегетативных частей, которые содержат большое количество клетчатки. Так как организмы большинства животных НЕ продуцируют целлюлозолитических ферментов, то расщепление клетчатки осуществляется симбиотическими бактериями и некоторыми простейшими кишечного тракта.
Среди млекопитающих переработка клетчатки особенно специфична у жвачных животных. Широко известный пример такой специфики – многокамерный желудок, состоящий из рубца и сычуга. У других млекопитающих основным местом переработки клетчатки является толстый кишечник, особенно его слепой отдел (слепые кишки или выросты). Химизм этой обработки сходен с пищеварением в рубце. У зайцеобразных и многих грызунов в связи с таким типом пищеварения вырабатывается два типа помета:
- первичный (который поглощается слепым отделом кишечника для повторной переработки) и
- вторичный (более сухой, который выводится из организма).
Использование первичного помета способствует более полной утилизации содержащегося в нем белка (включая бактериальные клетки).
Аналогичным образом в области толстого кишечника и его слепых отростков идет переваривание клетчатки у птиц, рацион которых частично или полностью состоит из растительных тканей: у врановых, гусеобразных, куриных. В максимальной мере такой тип пищеварения развит у тетеревиных птиц, т.е. именно у той группы, которые специализированны к переработке грубых веточных кормов. Например, длина только одной слепой кишки у взрослого глухаря-самца достигает почти одного метра. В летнее время, когда эти птицы имеют возможность потреблять более калорийные корма, длина и диаметр слепых кишок сокращается. Процесс перехода этих птиц с летних на зимние корма и обратно происходит постепенно и, вероятно, имеет «дружественный характер» по отношению к микрофлоре и бактериофауне, населяющей слепой отдел. У некоторых молодых птиц, впервые переходящих на зимние смолосодержащие корма (хвою, побеги сосны, кедра, ели, лиственницы), порой отмечается расстройство желудка и даже диарея.
Переваривание клетчатки с участием бактерий и простейших – широко распространенное явление и среди беспозвоночных животных. У низших термитов эта функция осуществляется в основном с помощью простейших (жгутиковых), у высших термитов и других групп большее значение имеют бактерии. Среди беспозвоночных имеются и формы, которые продуцируют для расщепления клетчатки собственные ферменты.
Для животных-фитофагов сбор корма это совсем не легкий, примитивный процесс, т.к. выбор мест кормления определяется не только расположением участков с изобильной пищей или риском хищничества. Большинство растений в процессе своей эволюции выработали специфические приспособления к поеданию фитофагами. В том числе и химическую защиту, которая сводится к выработке растениями соединений, которые препятствующих либо поеданию их органов животными, либо ухудшают переваривание корма животными (различные фенолы, танины, терпены и др. дубильные вещества). Например, фрагменты ели (практически все кроме семян) почти не поедаются животными, напротив, почти все фрагменты осины и ивы употребляются в пищу множеством фитофагов разных систематических групп. Эти защитные вещества являются вторичными метаболитами, и их концентрация в растениях существенно варьирует по годам, по сезонам, по местам их локализации в органах растений, по возрасту самих растений и т.д. Поэтому не случайно, что в последнее время процесс питания диких млекопитающих-фитофагов нередко рассматривается как постоянное пищевое «тестирование» животными конкретных видов и форм растений. Один из классических примеров «химического» предпочтения конкретных кормовых деревьев у птиц-фитофагов демонстрирует глухарь. Зимой он предпочитает питаться хвоей поврежденных сосен, у которых (по сравнению с другими соснами) в результате изменения метаболизма в хвое накапливается больше сахаров.
Консументы 2го порядка. Этот уровень объединяет животных с плотоядным типом питания – зоофагов.
Обычно в этой группе рассматривают всех хищников, т.к. их специфические черты практически не зависят от того, является ли жертва фитофагом или плотоядна. Таким образом, понятие «хищничество» в данном случае используется в широком экологическом смысле – как плотоядный тип питания, при котором животное-зоофаг, нападая на жертву, убивает ее. Но, строго говоря, консументами 2го порядка следует считать только хищников, питающихся растительноядными животными и, соответственно, представляющих второй этап трансформации органического вещества в цепях питания.
Химические вещества, из которых строятся ткани живого организма, довольно однородны. Поэтому пищевая трансформация вещества при переходе от фитофагов к консументам 2го порядка не имеет столь принципиального характера, как преобразование растительных тканей в животные.
При более детальном подходе уровень консументов 2го порядка следует разделять на подуровни, соответствующие направлению потоков вещества и энергии. Например, в трофической цепи «злаки – кузнечики – лягушки – змеи – орлы» лягушки, змеи и орлы составляют последовательные подуровни консументов 2го порядка.
Зоофаги, также как и фитофаги характеризуются своими специфическими приспособлениями к характеру питания.
Морфологически это выражается в специфических приспособлениях к характеру питания. Они имеют специальные приспособления для умерщвления жертв: когти, мощные клыки или клювы. Хотя есть и исключения: обыкновенный ёж не имеет клыков, т.е. зубов, приспособленных для убийства жертв. Он не способен умертвить крупную жертву (например, лягушку) и поедает ее живой. В этом случае жертва гибнет от болевого шока, реже от потери крови. Мне приходилось наблюдать, как серые вороны поедали кишечник еще живого сизого голубя. Чаще такое кормовое поведение распространено среди животных со смешанным типом питания, которые способны потреблять как растительную, так и животную пищу.
При питании животными, имеющими плотные защитные покровы, у хищников развиваются приспособления для их разрушения. Например:
- специализированные зубы у скатов («терка»), которыми они раздавливают панцири иглокожих моллюсков;
- широкие коренные зубы выдр-каланов снабжены прочными округлыми бугорками, которые помогают этим животным справляться с морскими ежами.
Для врановых и крупных чаек известно, что они способны схватив твердую добычу (моллюска, краба и др.) взлетать и с высоты бросать ее на землю. Такой прием они повторяют до тех пор, пока раковина или панцирь не расколется. Есть сообщения о том, что аналогичным образом поступают некоторые настоящие хищные птицы с черепахами (стервятник) или с костями крупных животных (бородач).
Адаптации зофагов на физиологическом уровне выражаются в основном в специфичности действия ферментов, «настроенных» на переваривание пищи животного происхождения. В опытах с собаками показано, что такая «настройка» может меняться: предварительное кормление растительными белками (хлебом) усиливает активность пепсина к этим белкам и уменьшает перевариваемость мяса.
Для многих фитофагов (т.е. для потенциальных жертв плотоядных животных) характерно наличие плотных хитиновых и кутикулярных оболочек, которые обычно не перевариваются большинством хищников. Это когти, копыта, рога, шерсть, перья. В основном они либо не поедаются хищниками, либо выводятся из их организмов в виде погадок или непереваренных остатков с экскрементами.
Однако у некоторых животных в желудке обнаруживаются ферменты, разлагающие хитин, причем у некоторых продукция этих ферментов связана с деятельностью продуктов-симбионтов.
Известны и специфические приспособления к питанию:
- хищные моллюски, питающиеся особями того же типа, «просверливают» их раковины с помощью концентрированных минеральных кислот, продуцируемых специальными железами;
- у кровососущих летучих мышей – вампиров – в секрете слюнных желез содержатся антикоагулянты, которые препятствуют свертыванию крови. Выделение антикоагулянтов характерно для многих пиявок и кровососущих членистоногих.
Говоря о типичных хищниках, следует отметить их поведенческие (или этологические) адаптации к кормодобыванию. Эти адаптации весьма и весьма разнообразны. Можно выделить следующие типы поиска и добывания корма хищниками.
- Поисковое обследование трофотопа (трофотоп – участок территории с потенциальными кормовыми объектами). В процессе такого обследования хищник находит жертву и, приблизившись к ней, добывает ее. Нередко такое обследование включает быстрое перемещение хищника из одной среды в другую. Например, горностай или ласка разыскивают корм и на поверхности снега, и под ним, и в каменистых россыпях, и под завалами деревьев. Куница обследует и поверхность почвы, и кроны деревьев.
- Охота из засады, когда хищник располагается в укромном месте, не редко на пути постоянных маршрутов своих жертв, или в местах с хорошим обзором. В таком случае при добыче жертв нередко используется фактор внезапности. Этим приемом пользуются самые разные хищники, в том числе и виды, для которых обычно активное поисковое поведение. Например, волк в Якутии ловит зайцев на тропах, которые зайцы набивают в глубоком снегу. Росомаха нередко нападает на молодняк копытных из крон деревьев, расположенных над тропами.
- Облавная охота и охота загоном. Используется хищниками, ведущими групповой образ жизни. В перовом случае (облава) хищники движутся цепью, «выталкивая» жертв на своих сородичей (нередко облавы используются волком при охоте на зайцев). При охоте загоном хищники, преследуя жертву, «направляют» ее движение к месту, где затаились их соплеменники (например, львы, волки). В горах хищники иногда загоняют жертв на обрывы, после чего поедают сорвавшихся с них особей. Описан случай, когда волк, выманивая собак чабанов, подводил собак к месту засады, где прятались другие волки.
Это лишь некоторые примеры. В действительности охотничье поведение хищников весьма разнообразно. И чем разнообразнее жертвы хищника, тем выше доля паратипической изменчивости в его генотипическом поведенческом стереотипе. Один из наиболее ярких примеров – подводная атака ястреба-тетеревятника.
Консументы 3го порядка. Выделение этого уровня весьма условно. Сюда относят животных с плотоядным типом питания, чаще всего имея в виду паразитов животных и «сверхпаразитов», хозяева которых сами ведут паразитический образ жизни. Явление сверхпаразитизма побуждает делить и этот трофический уровень на подуровни.
Но, строго говоря, сюда относятся все животные, поедающие плотоядных животных, тогда как паразиты растительноядных форм должны рассматриваться как консументы 2го порядка.
Деление биоценоза на трофические уровни – это лишь общая схема. Действительность сложнее. В частности существует много видов со смешанным питанием, и их можно относить к разным трофическим уровням.
Переход биомассы с нижнего трофического уровня на высший связан с потерями вещества и энергии. Считается, что в среднем около 10% биомассы и связанной в ней энергии переходит с каждого предыдущего уровня на следующий. По этой причине суммарная биомасса, продукция и энергия, а часто и численность особей прогрессивно уменьшаются по мере восхождения по трофическим уровням. Эта закономерность сформулирована Ч.Элтоном в виде правила экологических пирамид (рис. 3-1) и выступает как главный ограничитель длины пищевых цепей.
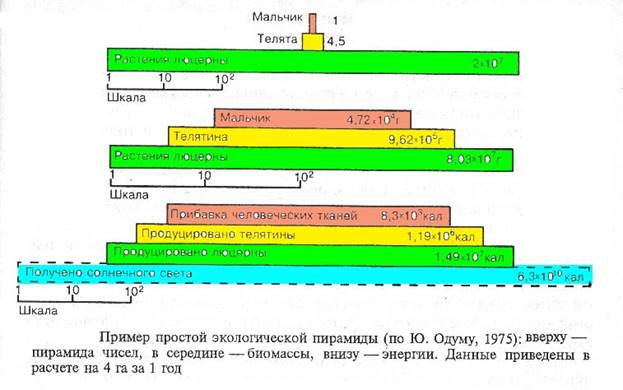
Рисунок 3-1.
Редуценты и детритофаги
Когда растения и животные погибают они НЕ перестают быть биологическими объектами, состоящими из органических соединений. Эти соединения продолжают представлять трофический интерес для других живых организмов и, соответственно, тела погибших становятся ресурсом для других организмов. Конечно, в известном смысле все консументы существуют за счет мертвого материала (зеленый лист в пищеводе становится мертвым, как только начинается переваривание). Однако во многих случаях организм, погибший НЕ в результате хищничества, также продолжает сохранять определенную трофическую ценность для других организмов (например, засохшая трава, опавшая листва, птица, погибшая при столкновении с проводами ЛЭП и др.).
На данном занятии мы будем говорить о редуцентах (к которым относятся бактерии и грибы) и детритофагах, т.е. о животных разных систематических групп, которые потребляют мертвый материал (черви, улитки, мокрицы, млекопитающие, птицы и др.). Совокупность редуцентов и детритофагов называется деструкторами. Деструкторы способны разлагать (разрушать) крупные органические молекулы на составные части. Их основная функция в биоценозе – минерализация органического вещества до простейших соединений: СО2, воды, различных солей и других неорганических соединений. При этом некоторые химические элементы на некоторое время включаются в состав тела организмов деструкторов, а энергия, которая высвобождается при разрушении органического вещества, служит источником их жизни.
Включение неорганических биогенных элементов (фосфора, азота, калия и др.) в состав органического вещества называется фиксацией этих элементов. Она происходит в процессе роста зеленых растений. Но, очевидно, что должен быть и обратный процесс. И он действительно существует. Это – разложение, высвобождение энергии и минерализация химических веществ, т.е. их превращение из органической в неорганическую форму. Разложение – это постепенное разрушение мертвого органического материала до неорганических соединений, которое осуществляется с помощью и физических факторов, и биологических агентов. Последними, т.е. биологическими агентами, и являются деструкторы.
Их основное отличие от растительноядных, плотоядных и паразитов заключается в том, что хищники и паразиты непосредственно влияют на скорость продуцирования своих пищевых ресурсов. Деструкторы НЕ контролируют скорость, с которой их ресурсы становятся доступными или возобновляются. Они полностью зависят от скорости, с которой какой-нибудь иной фактор (старение, болезни, конфликты, обвалы, оползни и др.) обеспечивает их пищевыми ресурсами. Модели, которые имитируют деятельность редуцентов и детритофагов показывают, что их взаимодействия с «донором» (т.е. с их пищей) оказываются весьма устойчивыми, и эта устойчивость НЕ зависит от увеличения видового разнообразия и сложности пищевой цепи, эта устойчивость даже возрастает. Такая ситуация совершенно противоположна взаимоотношения хищник-жертва, которые описываются моделью Лотки-Вольтерры. В отношениях деструкторов с их пищевыми ресурсами НЕТ обратной связи.
Объектами питания деструкторов служат самые разнообразные органы и покровы. В первую очередь это останки растений: отмершие части трав, кустарников и деревьев (включая древесные стволы, ветви, стебли, листья, почечные чешуйки и др.); это тела павших животных, в том числе и остатки трапезы хищников (включая фрагменты их скелетов), а также регулярно возобновляемые органы и покровы: например, плоды и семена растений, личиночные покровы членисоногих, старая кожа змей, волосы млекопитающих, перья, роговые оболочки птиц и млекопитающих, рога млекопитающих и др. Кроме того, у высших растений постоянно разрушаются клетки на корневых чехликах, а клетки корня отмирают по мере прорастания корней сквозь почву. Все эти отмершие образования также становятся добычей деструкторов.
Наконец, экскременты животных, независимо от того выделены ли они детритофагами, фитофагами, истинными хищниками или паразитами, также становятся кормовым ресурсом для деструкторов. Такое разнообразие органических остатков требует и разнообразных механизмов их утилизации.
Всю эту совокупность отмерших органических материалов можно несколько условно назвать термином детрит. Хотя обычно это понятие употребляется в более узких смыслах. Например, к детриту не причисляются трупы погибших животных.
Среди деструкторов выделяют две специализированные группы. Некрофаги – питающиеся в основном погибших трупами животных (грфы, гиены и др.); многие хищники часто используют некрофагию, особенно в условиях сложной кормовой обстановки. Другая специализированная группа – копрофаги – организмы, питающиеся преимущественно экскрементами животных (в основном беспозвоночные, но факультативная копрофагия отмечена и для целого ряда высших позвоночных).
Дата добавления: 2021-04-21; просмотров: 1977;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
