Гормон роста, пролактин
Гормон роста синтезируется в соматотрофных клетках, наиболее многочисленных в передней доле гипофиза. Содержание гормона роста составляет 5-16 мг в 1 г ткани железы, в то время как количество других гормонов гипофиза ис-
числяется в мкг/г. T1/2 гормона в плазме крови составляет около 50 мин.
Гормон роста у всех видов млекопитающих представляет собой одноцепочечный пептид с молекулярной массой 22 кД, состоящий из 191 аминокислотного остатка и имеющий 2 внутримолекулярные дисульфидные связи (рис. 11-10).
Гормон роста образуется из прогормона с молекулярной массой 28 кД, не обладающего гормональной активностью. Уровень гормона роста в плазме крови не превышает 3 нг/мл. Секреция гормона роста носит пульсирующий характер с интервалами в 20-30 мин. Один из самых больших пиков отмечается вскоре после засыпания.
Под влиянием различных стимулов (стресс, физические упражнения, гипогликемия, голодание, белковая пища, аминокислота аргинин) даже у нерастущих взрослых людей уровень гормона роста в крови может возрастать до 30-100 нг/мл.
Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. Основной стимулирующий эффект оказывает соматолиберин, основной тормозящий - ги-поталамический соматостатин.
Рецепторы гормона роста находятся в плазматической мембране клеток печени, жировой ткани, яичках, жёлтом теле, скелетных мышцах, хрящевой ткани, мозге, лёгких, поджелудочной железе, кишечнике, сердце, почках, лимфоцитах. Рецептор гормона роста - белок с одним внутримембранным доменом и молекулярной массой 70 кД. Связывание рецептора с гормоном роста вызывает димеризацию 2 рецепторов, что приводит к активации связанных с рецептором Янус-киназ и фосфорилированию Янус-киназ и рецептора по остаткам тирозина. Активация рецептора гормона роста сопровождается повышением активности тирозинкиназ и фосфо-липазы С с последующим повышением уровня ДАГ и ИФ3 и активацией протеинкиназы С (см. раздел 5).
Первичные эффекты гормона роста кратковре-менны и инсулиноподобны. Они проявляются в основном в отношении обмена жиров и углеводов. В жировой ткани усиливается потребление
глюкозы и липогенез, вследствие чего происходит снижение концентрации глюкозы в крови. Однако в дальнейшем проявляются более медленные (в основном, противоположные инсулину) эффекты: усиливается липолиз в жировой ткани, увеличивается концентрация жирных кислот в крови, а в случае недостаточности инсулина увеличивается содержание кетоновых тел в крови. Энергия, образующаяся при повышенном распаде жиров, используется на анаболические процессы. В то же время использование глюкозы жировыми и мышечными клетками снижается, а в печени ускоряется глюконеогенез, следствием чего может быть гипергликемия, особенно при недостатке инсулина (рис. 11-11).
Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных с ростом и развитием организма. Под влиянием гормона роста усиливаются транспорт аминокислот в клетки мышц, синтез белка в костях, хрящах, мышцах, печени и других внутренних органах, увеличивается общее количество РНК, ДНК и общее число клеток.
Влияние гормона роста на рост скелета и мягких тканей требует участия веществ, которые синтезируются в ответ на взаимодействие
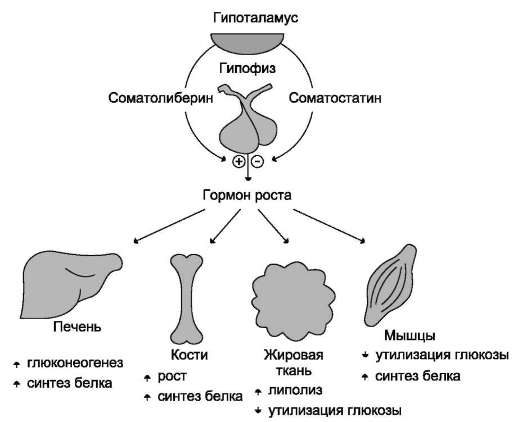
Рис. 11-11. Биологическое действие гормона роста.
гормона роста с рецепторами плазматической мембраны клеток различных тканей, в основном печени, и носят название соматомединов. Поскольку эти молекулы отличаются высокой гомологичностью друг к другу, а также к проинсулину и обладают инсулиноподобной активностью и мощным ростстимулирующим действием, они называются инсулиноподобны-ми факторами роста (ИФР-1, или соматомедин С; ИФР-2, или соматомедин А). ИФР-1 - од-ноцепочечный полипептид основного характера, содержащий 70 аминокислотных остатков, а полипептид ИФР-2 носит кислотный характер и состоит из 67 аминокислотных остатков. В крови примерно 95% соматомединов циркулирует в комплексе с белками. Синтез ИФР-1 в большей степени зависит от концентрации гормона роста в крови, чем синтез ИФР-2. В то же время ИФР-1, образующийся в печени, ингибирует синтез и секрецию гормона роста
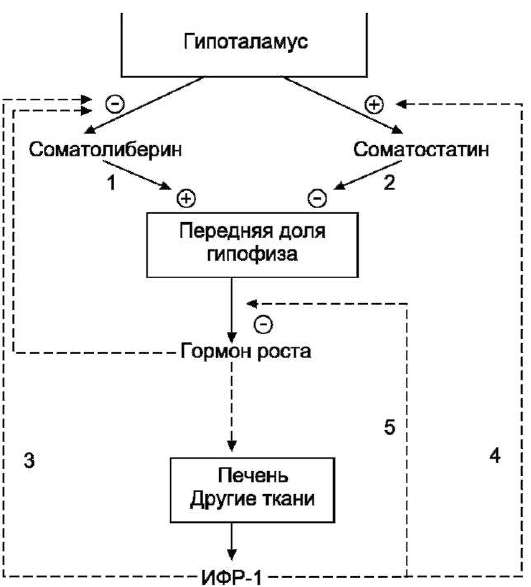
Рис. 11-12. Регуляция секреции гормона роста.
Соматолиберин стимулирует (1), а соматостатин ингибирует (2) освобождение гормона роста (ГР) из передней доли гипофиза. ИФР-1 ингибирует секрецию соматолиберина (3) и стимулирует секрецию сома-тостатина (4). ИФР-1 ингибирует секрецию гормона роста также на уровне гипофиза (5).
по механизму ретроингибирования, действуя на уровне гипофиза и гипоталамуса (рис. 11-12).
Инсулиноподобные факторы роста оказывают своё действие различными путями: эндокринным, паракринным и аутокринным (рис. 11-13).
Подобно рецептору инсулина, рецептор ИФР-1 обладает тирозинкиназной активностью и инициирует каскад реакций фосфорилирования других белков, участвующих в различных внутриклеточных процессах, включая активацию транскрипции генов. В большинстве случаев ИФР-1, как и инсулин, инициирует клеточное развитие, однако при значительно меньших, почти физиологических концентрациях. Это указывает на то, что инсулиноподобные факторы роста более активны в отношении их действия на рост и развитие клеток.
Под влиянием гормона роста увеличивается ширина и толщина костей, и одновременно с этим ускоряется рост других тканей, включая соединительную ткань, мышцы и внутренние органы.
Пролактинсинтезируется лактотрофными клетками передней доли гипофиза в виде про-гормона с молекулярной массой 40 кД. Число этих клеток резко возрастает при беременности под влиянием эстрогенов. Пролактин близок по химическому строению гормону роста. Он состоит из 199 аминокислотных остатков, образующих одну полипептидную цепь с тремя дисульфидными связями. 35% аминокислотной последовательности пролактина идентично последовательностям гормона роста. Оба гормона имеют общие антигенные детерминанты, сходное строение рецепторов и пути трансдукции сигналов в клетки.
Рецепторы пролактина присутствуют в клетках многих тканей: в печени, почках, надпочечниках, яичках, яичниках, матке и других тканях.
Основная физиологическая функция про-лактина - стимуляция лактации. Пролактин индуцирует синтез α-лактальбумина и казеина, активирует синтез фосфолипидов и ТАГ.
На процессы роста пролактин влияет в значительно меньшей степени, чем гормон роста.
У мужчин пролактин повышает чувствительность клеток Лейдига к лютеинизирующему гормону, поддерживая таким образом необходимый уровень синтеза тестостерона; в почках пролак-
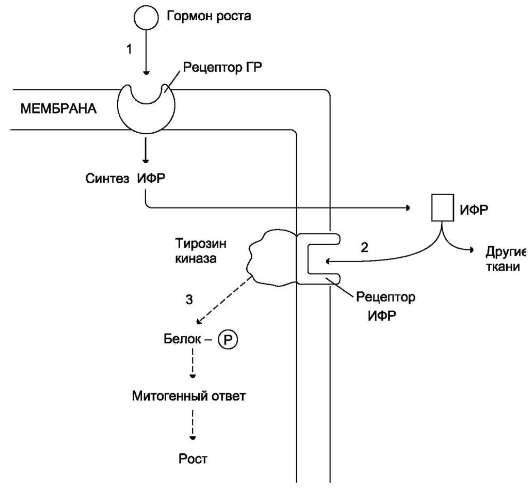
Рис. 11-13. Действие гормона роста через ИФР.Гормон роста взаимодействует с рецептором плазматической мембраны клеток, стимулируя синтез ИФР (1). ИФР, в свою очередь, взаимодействуют со специфическими рецепторами клеток той же или других тканей (2) и стимулируют фосфорилирование белков, участвующих в митозе и росте (3).
тин снижает экскрецию воды, влияет на реабсор-бцию ионов Na+ и К+; пролактин также повышает гуморальный и клеточный иммунитет.
Синтез и секрецию пролактина стимулируют тиреолиберин, серотонин, окситоцин, ацетил-холин, ингибирующий эффект оказывает дофамин.
Подобно большинству гормонов, пролактин секретируется в кровь эпизодически с интервалами 30-90 мин. Максимум секреции отмечается через 6-8 ч после начала сна. Концентрация пролактина в плазме крови женщин составляет 8-10 нг/мл, а мужчин - 5-8 нг/мл. T1/2 пролактина составляет 15-20 мин.
Плацента продуцирует гормон (плацентарный лактоген), гомологичный по аминокислотному составу гормону роста и пролактину. Все 3 гормона имеют общие антигенные детерминанты
и обладают рост-стимулирующей и лактогенной активностью. Существует гипотеза, согласно которой гены этих гормонов возникли в результате дупликации одного гена-предшественника.
Дата добавления: 2017-01-08; просмотров: 5416;
Поиск по сайту
Узнать еще
- Iii. строение, биосинтез и биологическое действие гормонов
- IV. Взаимодействие гормона с клеткой-мишенью
- Б. Функция кальцитонина. В. Функция парагормона.
- Биотехнология получения инсулина, гормона роста и интерферона
- Биотрансформации стероидных гормонов
- В зависимости от пола, возраста и роста, кг
- В. ИЗМЕНЕНИЕ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ ГОЛОДАНИИ
- В. КЛАССИФИКАЦИЯ И НОМЕНКЛАТУРА ГОРМОНОВ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
