Механизм действия и патогенез интоксикации
ФОС относятся к антихолинэстеразным ядам. Действие их на организм объясняется способностью угнетать (ингибировать) ацетилхолинэстеразу и нарушать гидролиз ацетилхолина, осуществляющего передачу нервного импульса в холинергических синапсах.
Структура, в которой осуществляется функциональный контакт между двумя клетками, называется синапсом (рис. 9). Согласно биохимической теории, передача нервного импульса в синапсах происходит посредством медиаторов (переносчиков). В холинергических нейронах медиатором служит ацетилхолин, в адренергических - норадреналин. В состав синапса входит концевое утолщение нерва, вакуоли, содержащие связанный ацетилхолин, пресинаптическая мембрана с утолщениями и отверстиями, синаптическая щель шириной 20-50 нм, постсинаптическая мембрана с холинорецепторами.
В состоянии покоя ацетилхолин находится в связанном состоянии, постсинаптическая мембрана в состоянии электрической поляризации: внутри клетки содержится больше отрицательно заряженных анионов, снаружи ионов K и Ca, создается мембранный потенциал покоя около 50-90 мВ.
Под действием нервного импульса ацетилхолин высвобождается из вакуолей и выходит в синаптическую щель, где он сразу же вступает во взаимодействие с холинорецептором постсинаптической мембраны, передавая импульс на другой нейрон или эффекторный орган, и обеспечивает выполнение определенных функций органа. При этом одновременно происходит деполяризация постсинаптической мембраны, вследствие повышения ее проницаемости и диффузии ионов натрия внутрь.
После передачи импульса ацетилхолин почти мгновенно (за 0,002 с) гидролизуется под действием фермента ацетилхолинэстеразы на уксусную кислоту и холин. Одновременно происходит поляризация мембраны за счет выхода ионов Na и готовность синапса к проведению следующего импульса.
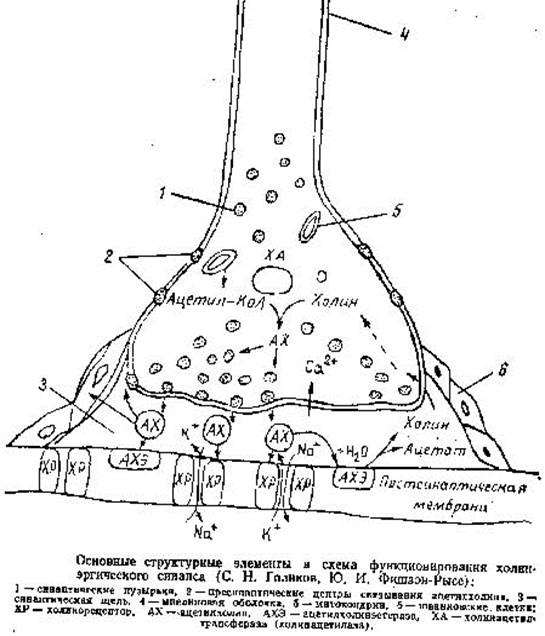
Рисунок 9.
Затем ацетилхолин синтезируется в пресинаптическом окончании путем переноса ацетильной группы ацетилкоэнзима-А на холин с помощью холинацетилтрансферазы при участии АТФ.
Холинэстераза - фермент, содержащийся в организме всех животных, птиц, рыб, насекомых, имеющих нервную систему. Установлено, что на ее активном центре имеются два активных участка. Анионный отрицательно заряженный участок образован СОО-радикалом, притягивает положительно заряженную головку ацетилхолина. Эстеразный участок имеет более сложную структуру, образован гидроксилом аминокислоты серина и имидазольным кольцом гистидина (акцептором и донором протона), обеспечивает связь со сложноэфирной группой ацетилхолина и ее разложение.
Холинорецепторы представляют собой белковолипидные комплексы, реагирующие с ацетилхолином, воспринимающие нервный импульс и передающие действие на следующий нейрон или эффекторный орган. В холинорецепторах также имеются два активных центра: анионный, притягивающий положительно заряженную головку ацетилхолина, и эстерофильный, реагирующий со сложноэфирной связью ацетилхолина (но не разлагающий ее). При этом происходит деполяризация постсинаптической мембраны, вследствие выхода из клетки ионов К и проникновения ионов Na.
Фармакологическими методами установлено два вида холинорецепторов. Мускариночувствительные М-холинорецепторы находятся в синапсах ЦНС, постганглионарных нервных окончаниях парасимпатической нервной системы и синаптических нервных окончаниях потовых желез, они чувствительны к мускарину, блокируются М-холинолитиками (атропином). Никотинчувствительные Н-холинорецепторы содержатся в парасимпатических и симпатических ганглиях, где они возбуждаются малыми дозами никотина и угнетаются большими дозами никотина и ганглиоблокаторами, а также в мионевральных синапсах, поперечно-полосатой мускулатуре, в ЦНС.
Дата добавления: 2016-11-04; просмотров: 1791;
Поиск по сайту
Узнать еще
- CIH — газораспределительный механизм с распределительным валом в головке блока цилиндров
- II группа действий. ВЗАИМОДЕЙСТВИЯ
- II. ОБУЧЕНИЕ ПРОСТЫМ БОЕВЫМ ДЕЙСТВИЯМ
- II. Получение вращающегося магнитного поля и принцип действия АД.
- II. Принцип действия и режимы работы синхронной машины
- II. Электрическая схема и принцип действия.
- III. БОЕВЫЕ ДЕЙСТВИЯ
- III. Механизмы регуляции количества ферментов

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
