Надорганизменные биосистемы. Популяции
Организмы одного вида в природе всегда представлены не по отдельности, а определенным образом организованными совокупностями - популяциями*. Популяции могут быть монолитными или состоять из группировок субпопуляционного уровня - семей, кланов, стад, стай и т.п. Объединение организмов одного вида в популяцию выявляет качественно новые свойства. Решающее значение приобретают численность и плотность организмов, их пространственное размещение, половой и возрастной состав, характер взаимоотношений между особями, размежевание или контакты с другими популяциями этого вида и т.д. По сравнению с временем жизни отдельного организма популяция может существовать очень долго.
Вместе с тем популяция обладает и чертами сходства с организмом как биосистемой, так как имеет определенную структуру, целостность, генетическую программу самовоспроизведения, способность к авторегуляции и адаптации, свое коллективное материально-энергетическое хозяйство. Популяции являются реальными единицами биомониторинга, эксплуатации и охраны природных экосистем. Взаимодействие людей с видами организмов, находящихся в среде, в природном окружении или под хозяйственным контролем человека, опосредуется, как правило, через популяции. Это могут быть штаммы болезнетворных или полезных микроорганизмов, сорта возделываемых растений, породы разводимых животных, естественные популяции промысловых рыб и т.п. Не менее важно и то, что многие закономерности популяционной экологии относятся к популяциям человека.
Структуре популяция. Различают половую, возрастную, генетическую, пространственную и экологическую структуру популяций.
Половая структура популяции - это соотношение в ней особей разного пола. Существенное значение она имеет для тех форм, у которых четко выражена полная бисексуальность - преимущественно для членистоногих и позвоночных животных. У большинства из них соотношение полов определяется различием хромосомных наборов мужских и женских особей. Такое двухфакторное хромосомное определение пола обеспечивает равную численность полов (первичное соотношение полов). Но у некоторых животных наблюдается не двухфакторное, а трех- и более факторное генетическое определение пола. Это приводит к более сложной половой структуре популяций и заметному отклонению в соотношении полов (чаще в сторону преобладания женских особей).
В ряде случаев соотношение полов определяется не генетическими, а физиологическими, гормональными факторами и условиями среды, действующими во время и после оплодотворения (вторичное соотношение полов). Например, у многих рептилий, а также у муравьев и термитов формирование пола существенно зависит от температуры эмбрионального развития. Наконец, известны примеры, когда изменение экологических условий по-разному влияет на смертность самцов и самок. Это приводит к колебаниям их соотношения от года к году и к тому, что в разных популяциях одного вида (например, у некоторых полевок) соотношение самцов и самок может оказаться различным (третичное соотношение полов}.
Возрастная структура популяции - это соотношение в составе популяции особей разного возраста, представляющих один или разные приплоды одного или нескольких поколений. Поколение может состоять из особей одного приплода и из особей разных приплодов (например у мелких млекопитающих). Возрастная структура популяции отражает интенсивность размножения, уровень смертности, скорость смены поколений. Для всех популяций в природе справедливо правило стабильности половозрастной структуры: любая популяция в соответствии с условиями ее существования стремится к определенному оптимальному распределению особей по полу и возрасту.
Генетическая структура популяции определяется изменчивостью и разнообразием генотипов, частотами вариаций отдельных генов - аллелей, а также разделением популяции на группы генетически близких особей, между которыми при скрещивании происходит постоянный обмен аллелями. Для каждой популяции характерен также определенный уровень фенотипического полиморфизма, т.е. разнообразия признаков организма, находящихся под совместным контролем генов и экологических факторов. Один и тот же генотип в разных условиях способен привести к появлению различающихся фенотипов. Разнообразие генотипов зависит от размера популяции и внешних факторов, влияющих на ее структуру. Чем выше генетическая разнородность популяции, тем больше ее экологическая пластичность - возможность приспосабливаться к меняющимся условиям среды. В небольших изолированных и стабильных популяциях закономерно возрастает частота близкородственного скрещивания, что уменьшает генетическое разнообразие и увеличивает угрозу вымирания.
Пространственная структура популяции - это характер размещения и распределения отдельных членов популяции и их группировок на популяционной территории (ареале). В популяции реализуется принцип территориальности: все особи и их группы обладают индивидуальным и групповым пространством, возникающим в результате активного физико-химического или поведенческого разобщения. Оно часто сочетается с агрегацией, группировкой особей, которое усиливает конкуренцию между индивидами, но способствует выживанию группы в целом. Следовательно, как перенаселенность, так и недонаселенность, препятствующая агрегации, могут служить лимитирующими факторами. Так образуются стаи, стада, колонии и другие объединения особей, благодаря чему достигаются различные защитные эффекты. Пространственная структура популяций заметно различается у оседлых и кочующих или мигрирующих животных.
Экологическая структура популяции - это разделенность всякой популяции на группы особей, по-разному взаимодействующие с факторами среды. Легко выявляются группировки по питанию, так как особи разного пола и возраста обладают различным пищевым предпочтением. Половозрастные группировки связаны также с пространственным распределением. Разные члены популяции отличаются друг от друга по ориентировочному поведению и по двигательной активности; у многих животных хорошо выражены различия реакций избегания опасности или оптимизационного поиска. Часто наблюдается распределение функций («разделение труда») при охоте на добычу, при уходе за потомством и т.п. Наличие мигрирующих и немигрирующих групп особей накладывает отпечаток на ряд физиологических особенностей питания, полового поведения, групповой активности. Для всех популяций характерна, по-видимому, и фенологическая дифференциация: разные сроки начала и окончания сезонных циклов развития и поведения (диапауза, спячка, половая активность, линька, цветение, плодоношение, листопад и т.п.); наличие сезонных рас у насекомых, растений, проходных рыб.
Размер и динамика численности популяции. Для реализации нормальной структуры популяции она должна обладать некоторой минимальной численностью и плотностью, т.е. числом особей, приходящимся в среднем на единицу площади или объема. В зависимости от внешних и внутренних факторов численность и плотность популяций колеблется во времени - по годам, сезонам, от поколения к поколению.
Точная численность природных популяций может быть установлена только в случаях хорошей изоляции. Лучше всего в этом отношении изучены островные популяции некоторых видов животных. В разных популяциях растений и животных может быть и несколько десятков, и миллионы особей; они могут занимать территории и в несколько квадратных метров, и во многие тысячи квадратных километров. Размер популяционной территории связан с радиусом репродуктивной активности.
Если не принимать во внимание возможную миграцию, то численность популяции определяется соотношением рождаемости и смертности, на которые оказывают влияние внешние и внутренние популяционные факторы.
Потенциальная способность к размножению у многих организмов огромна. У простейших в благоприятных условиях промежуток между последовательными делениями может сокращаться до нескольких минут. Гриб склеропора, паразитирующий на кукурузе, порождает до 6 млрд спор на одно растение в день. Треска откладывает до 4 млн икринок в год, сельдь на протяжении жизни - от 8 до 75 млрд. У млекопитающих в одном помете от 1 (киты, слоны, приматы) до 20 особей (у серой крысы).
Высокая плодовитость компенсируется гибелью подавляющего большинства гамет и зачатков, а также родившихся особей из-за факторов сопротивления среды: недостатка пищи, действия неблагоприятных абиотических факторов, конкуренции, отклонений в развитии, болезней, паразитов, хищников, нехватка пространства, убежищ и т.п.
Изменения численности популяции в какой-то период определяются разностью относительных величин рождаемости и смертности. Ее называют биотическим, или репродукционным потенциалом г.
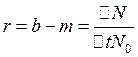 (3.1)
(3.1)
где b - относительная рождаемость (в процентах или в промилле),
т - относительная смертность,
DN - изменение численности популяции за время М.
N0 - исходная численность.
При отсутствии сопротивления среды наблюдается экспоненциальный рост популяции, так как прирост числа особей пропорционален уже имеющемуся их числу (рис. 3.1, А). Это выражается дифференциальным уравнением
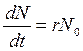 или иначе
или иначе 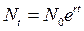 (3.2)
(3.2)
В популяции микроорганизмов, которая каждые два дня увеличивается в 10 раз, г = 1,15/сут. Для амбарного долгоносика, полевой мыши и человека г составляют соответственно 39,6; 4,5 и 0,02 в год; это означает удвоение популяции соответственно через 1 неделю, 8 недель и 35 лет. Между репродуктивным потенциалом и временем генерации у разных организмов существует четко выраженная обратная зависимость.
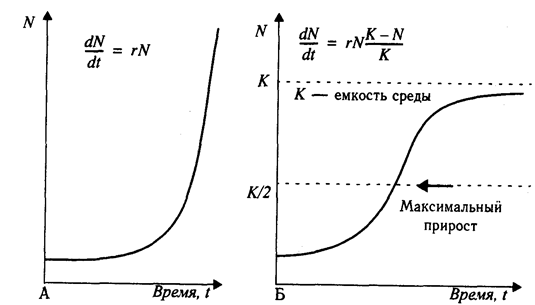
Рис. 3.1. Кривые роста численности популяций
А - экспоненциальная кривая роста при идеальных условиях среды;
Б - логистическая кривая роста в реальных естественных условиях при емкости среды, равной К.
В природных условиях рост популяции рано или поздно прекращается из-за сопротивления среды, которое тем больше, чем больше численность популяции. Поэтому реальная кривая роста принимает сигмовидную форму (рис. 3.1,Б), подчиняясь зависимости:
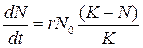 (3.3)
(3.3)
После начальной логарифмической фазы она асимптотически приближается к уровню максимальной численности и плотности насыщения, когда смертность равна рождаемости (b = т). К обычно обозначают как емкость среды.
Размер популяции поддерживается вблизи К различными способами. У видов, живущих в ненадежных местообитаниях с высоким сопротивлением (большими потерями от врагов, болезней, случайных колебаний климатических условий) или у паразитов (малые шансы найти хозяина), репродуктивный потенциал должен быть очень большим. Это так называемые г-стратеги - протисты, низшие растения, паразитические черви, многие рыбы. Напротив, виды, освоившие среду с малым сопротивлением, или виды с развитой заботой о потомстве, образующие семьи или стада, обходятся малым репродуктивным потенциалом. Это К-стратеги - орлы, киты, крупные копытные, приматы. Все же в большинстве случаев регуляция плотности популяции осуществляется ниже уровня емкости среды, не доводя до предельного использования ресурсов энергии и пространства.
В связи с изменениями условий среды численность популяций постоянно изменяется. Обычно колебания численности неупорядоченны и зависят от случайного сочетания многих факторов. Если сопротивление среды длительное время понижено, например благодаря благоприятным погодным и кормовым условиям, то у видов с перекрывающимися поколениями (прежде всего у г-стратегов), может наблюдаться быстрое размножение. Так происходят массовые вспышки численности у некоторых насекомых. Для разных видов существуют определенные амплитуды обычных колебаний численности. Для некоторых популяций (например, для мелких грызунов Субарктики) характерны относительно упорядоченные колебания численности с чередованием подъемов и спадов через определенные интервалы - от трех до десяти лет. Часто причиной такой цикличности являются взаимовлияния популяций хищника и жертвы.
При чрезмерной плотности популяции, перенаселенности ее территории, например, у мелких грызунов, наблюдаются признаки популяционного стресса и снижения жизнеспособности. Это выражается в более контрастном выделении доминантных и субдоминантных особей, проявлениях афессии, поведенческой дискриминации части особей, снижении плодовитости, ослаблении иммунитета и резком повышении вероятности эпизоотии. У некоторых видов популяционный стресс сопровождается массовой миграцией и расселением членов популяции.
Экосистемы
Популяция или часть популяции какого-либо вида растений или животных может входить в состав разных сообществ, где она сочетается с различными наборами представителей других видов. Многовидовые сообщества организмов в природе - биоценозы (см. стр. 31) - образованы не случайным сочетанием видов, а таким их подбором, такими количественными соотношениями и такой пространственно-временной организацией, которые обеспечивают непрерывный круговорот веществ и четко распределенные потоки энергии. Этим достигается устойчивость экосистем.
Состав м функциональная структура экосистемы. Каждая экосистема имеет собственное материально-энергетическое обеспечение и определенную функциональную структуру, основанную на пищевых (трофических) взаимоотношениях (рис. 3.2). Эта структура составлена несколькими группами организмов, каждая из которых выполняет определенную работу в круговороте веществ. Организмы, относящиеся к одному такому звену, образуют трофический уровень, а последовательные связи между трофическими уровнями образуют цепи питания, или трофические цепи. В экосистему входят организмы, различаемые по способу питания - автотрофы и гетеротрофы.
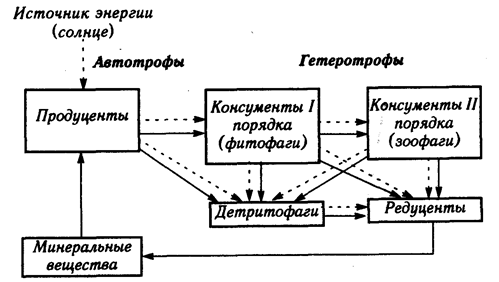
Рис. 3.2. Упрощенная схема переноса веществ и энергии в экосистеме
 - перенос веществ, - перенос энергии
- перенос веществ, - перенос энергии
Автотрофы (самопитающие) - организмы, образующие органическое вещество своего тела из неорганических веществ - в основном из углекислого газа и воды - посредством процессов фотосинтеза и хемосинтеза. Фотосинтез осуществляют фотоавтотрофы - все хлорофиллоносные (зеленые) растения и микроорганизмы. Хемосинтез наблюдается у некоторых почвенных и водных хемоавтотрофных бактерий, которые используют в качестве источника энергии не солнечный свет, а ферментативное окисление ряда веществ - водорода, серы, сероводорода, аммиака, железа.
Фотоавтотрофы (растения) составляют основную массу биоты и полностью отвечают за образование всего нового органического вещества в экосистеме, т.е. являются первичными производителями продукции - продуцентами экосистем. Синтезированная автотрофами новая биомасса органического вещества - это первичная продукция, а скорость ее образования - биологическая продуктивность экосистемы. Автотрофы образуют первый трофический уровень любой полночленной экосистемы.
Гетеротрофы (питающиеся другими) - организмы, потребляющие готовое органическое вещество других организмов и продуктов их жизнедеятельности. Это все животные, грибы и большая часть бактерий.
В отличие от автотрофов-продуцентов гетеротрофы выступают как потребители и деструкторы (разрушители) органических веществ. В зависимости от источников питания и участия в деструкции они подразделяются на консументов и редуцентов.
Консументы - потребители органического вещества организмов. К ним относятся:
§ консументы I порядка - растительноядные животные (фитофаги), питающиеся живыми растениями (тля, кузнечик, гусь, овца, олень, слон);
§ консументы II порядка - плотоядные животные (зоофаги), поедающие других животных, - различные хищники (хищные насекомые, насекомоядные и хищные птицы, хищные рептилии и звери), нападающие не только на фитофагов, но и других хищников. Существует немало животных со смешанным питанием, потребляющих и растительную и животную пищу - плотоядно-растительноядные и всеядные. Консументы I и II порядка занимают соответственно второй, третий, а иногда и следующий трофические уровни в экосистеме.
Особенную группу консументов составляют паразиты и симбиотрофы. И те и другие живут (по меньшей мере на протяжении части жизненного цикла) за счет веществ организма-хозяина. Это уже не только животные (черви, насекомые, клещи), но и различные микроорганизмы, а также некоторые грибы и растения. В отличие от паразитов, часто вызывающих заболевания, а иногда и гибель хозяина, симбиотрофы выполняют жизненно важные для хозяина трофические функции. Это мицелиальные грибы-микоризы, участвующие в корневом питании многих растений; клубеньковые бактерии бобовых, связывающие молекулярный азот; микробиальное население сложных желудков жвачных животных, повышающее перевариваемость и усвоение поедаемой растительной пищи.
Еще одну группу консументов образуют детритофаги, или сапрофаги - животные, питающиеся мертвым органическим веществом - остатками и продуктами жизнедеятельности растений и животных. Это различные черви, членистоногие (клещи, многоножки, личинки насекомых, жуки-копрофаги) и другие животные - все они выполняют функцию очищения экосистем. Детритофаги участвуют в образовании почвы, торфа, донных отложений водоемов.
Редуценты - бактерии и низшие грибы - завершают деструктивную работу консументов и сапрофагов, доводя разложение органики до ее полной минерализации и возвращая в среду экосистемы молекулярный азот, минеральные элементы и последние порции двуокиси углерода.
Все названные группы организмов в любой экосистеме тесно взаимодействуют между собой, согласуя потоки вещества и энергии. Их совместное функционирование не только поддерживает структуру и целостность биоценоза, но и оказывает существенное влияние на абиотические компоненты биотопа, формируя и поддерживая экологическую среду экосистемы.
Никакая часть экосистемы не может существовать без другой. Если по какой-либо причине происходит нарушение структуры экосистемы, исчезает группа организмов, вид, то по закону цепных реакций может сильно измениться или даже разрушиться все сообщество. Но часто бывает и так, что через какое-то время после исчезновения одного вида на его месте оказываются другие организмы, другой вид, но выполняющий сходную функцию в экосистеме. Эта закономерность называется правилом, замещения, или дублирования:
у каждого вида в экосистеме есть «дублер». Такую роль обычно выполняют виды менее специализированные и в то же время экологически более гибкие, адаптивные. Так, копытных в степи замещают грызуны; на мелководных озерах и болотах аистов и цапель замещают кулики и т.п. При этом решающую роль играет не систематическое положение, а близость экологических функций между замещаемыми и замещающими группами организмов.
Принципиальное различие между потоками вещества и энергии в экосистеме заключается в том, что биогенные элементы, составляющие органическое вещество, могут многократно участвовать в круговороте веществ, тогда как поток энергии однонаправлен и необратим. Каждая порция энергии используется однократно. В соответствии со вторым законом термодинамики на каждом этапе трансформации энергии значительная ее часть неизбежно рассеивается в виде теплоты.
Пищевые цепи и трофические уровни. Прослеживая пищевые взаимоотношения между членами биоценоза («кто кого и сколько поедает»), можно построить пищевые цепи питания различных организмов. Примером длинной пищевой цепи может служить последовательность обитателей арктического моря: «микроводоросли (фитопланктон) ® мелкие растительноядные ракообразные (зоопланктон) ® плотоядные планктонофаги (черви, ракообразные, моллюски, иглокожие) ® рыбы (возможны 2-3 звена последовательности хищных рыб) ® тюлени ® белый медведь». Цепи наземных экосистем обычно короче. Пищевая цепь, как правило, искусственно выделяется из реально существующей пищевой сети - сплетения многих цепей питания.
Благодаря определенной последовательности пищевых отношений различаются отдельные трофические уровни переноса веществ и энергии в экосистеме, связанные с питанием определенной группы организмов. Совокупности трофических уровней различных экосистем моделируются с помощью трофических пирамид чисел (численностей), биомасс и энергий (рис. 3.3). Обычные пирамиды чисел, т.е. отображение числа особей на каждом из трофических уровней данной экосистемы, для пастбищных (по Ю. Одуму. 1975) А - пирамида чисел, Б - пирамида биомасс, В - пирамида энергий. Данные приведены в расчете на 4 га за год; шкалы логарифмические цепей имеют широкое основание (большое число продуцентов, Р) и резкое сужение к конечным консументам (рис. 3.3, А). При этом числа «ступеней» различаются не менее, чем на 1 -3 порядка. Но это справедливо только для травяных сообществ - луговых или степных биоценозов. Картина резко искажается, если рассматривать лесное сообщество. На одном дереве могут кормиться тысячи фитофагов или на одном трофическом уровне оказываются такие разные фитофаги, как тля или слон.
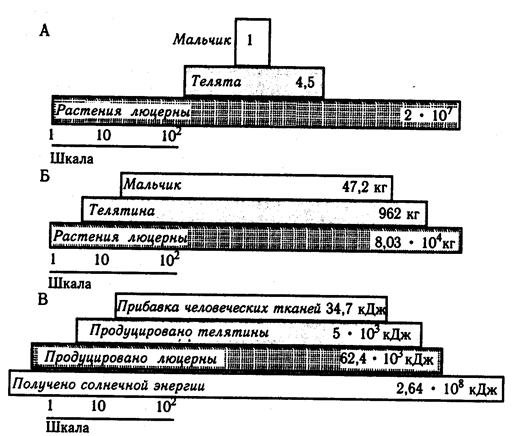
Рис. 3.3. Пример простой трофической пирамиды
Это искажение можно преодолеть с помощью пирамиды биомасс. В наземных экосистемах биомасса растений всегда существенно больше биомассы животных, а биомасса фитофагов всегда больше биомассы зоофагов (рис. 3.3, Б). Иначе выглядят пирамиды биомасс для водных, особенно морских экосистем: биомасса животных обычно намного больше биомассы растений. Эта «неправильность» обусловлена тем, что пирамидами биомасс не учитывается продолжительность существования поколений особей на разных трофических уровнях и скорость образования и выеданйя биомассы. Главным продуцентом морских экосистем является фитопланктон, имеющий большой репродуктивный потенциал и быструю смену поколений. В океане за год может смениться до 50 поколений фитопланктона. За то время, пока хищные рыбы (а тем более моржи и киты) накопят свою биомассу, сменится множество поколений фитопланктона, суммарная биомасса которых намного больше. Вот почему универсальным способом выражения трофической структуры экосистем являются пирамиды скоростей образования живого вещества, т.е. продуктивности. Их обычно называют пирамидами энергий, имея в вицу энергетическое выражение продукции, хотя правильнее было бы говорить о мощности.
Стабильность и развитие экосистем. В природных экосистемах происходят постоянные изменения состояния популяций организмов. Они вызываются разными причинами. Кратковременные - погодными условиями и биотическими воздействиями; сезонные (особенно в умеренных и высоких широтах) - большим годовым ходом температуры; от года к году - различными случайными сочетаниями абиотических и биотических факторов. Однако все эти колебания, как правило, более или менее регулярны и не выходят за границы устойчивости экосистемы - ее обычного размера, видового состава, биомассы, продуктивности, соответствующих географическим и климатическим условиям местности. Такое состояние экосистемы называется климаксным.
Климаксные сообщества характеризуются устойчивым динамическим равновесием между биотическими потенциалами входящих в сообщество популяций и сопротивлением среды. Постоянство важнейших экологических параметров обозначают как гомеостаз экосистемы. Устойчивость экосистемы тем. больше, чем больше она по размеру и чем богаче и разнообразнее ее видовой и популяционный состав.
Стремясь к поддержанию гомеостаза, экосистемы, тем не менее, способны к изменениям, к развитию, к переходу от более простых к более сложным формам. Масштабные изменения географической обстановки или типа ландшафта под влиянием природных катастроф или деятельности человека приводят к определенным последовательным изменениям состояния биогеоценозов местности - сукцессиям (от англ. succession - последовательность).
Различают первичную сукцессию - постепенное заселение организмами появившейся девственной суши, оголенной материнской породы (отступившее море или ледник, голые скалы и застывшая лава после вулканического извержения и т.п.). В этих случаях решающую роль играет процесс почвообразования. Начальное выветривание - разрушение и разрыхление поверхности минеральной основы под действием перепадов температуры и увлажнения - уже может быть использовано бактериями, лишайниками, а затем и редкой одноярусной пионерной растительностью. Ее появление, а с нею - симбиотрофов и мелких животных значительно ускоряет образование почвы и постепенное заселение территории сериями все более сложных растительных сообществ, все более крупными растениями и животными. Так система постепенно проходит все стадии развития до климаксного состояния.
Вторичные сукцессии имеют характер постепенного восстановления свойственного данной местности сообщества после нанесенных повреждений (последствий бури, пожара, вырубки, наводнения, выпаса скота, запуска полей). Возникшая в результате вторичной сукцессии климаксная система может существенно отличаться от первоначальной, если изменились некоторые характеристики ландшафта или климатические условия. Сукцессии происходят путем замещения одних видов другими и поэтому их нельзя приравнивать к реакциям гомеостаза.
Биомы. Наземные экосистемы, относящиеся к одной природно-климатической зоне имеют общую структуру доминирующей растительности и поэтому могут рассматриваться как единый большой биогеоценоз - биом. Биомы являются основными объектами экологической географии. Они различаются по климату, по многим особенностям флоры и фауны, по биологическому разнообразию, но в пределах каждого биома можно встретить множество сходных по приспособлению форм животных и растений, хотя происхождение их различно. В табл. 3.1 приведены ориентировочные данные о площади, биомассе и годовой продуктивности крупнейших биомов, охватывающих всю Землю. По ним можно судить не только о больших экологических различиях природных зон, но и о степени давления человека на природу планеты. Еще совсем недавно при составлении подобных таблиц и карт экологи пренебрегали участками суши, преобразованными человеком. Обращает на себя внимание, что 94% всей биомассы Земли сосредоточено в лесах, более половины - во влажных тропических лесах. Обрабатываемые земли, составившие 10% площади суши, дают такую же долю первичной продукции, уступая по продуктивности степям, которые вытеснены пашней.
Таблица 3.1
Площади, биомасса и продуктивность основных биомов Земли
| Основные биомы, категория земель | Площадь, млн км | Биомасса (сухое вещество) | Готовая продукция | ||
| т/га | млрд т. | т/га | млрд т. | ||
| Тундры и лесотундры | 4,2 | 2,1 | |||
| Таежные и горные хвойные леса | 12,8 | 11,5 | |||
| Лиственно-хвойные бореальные леса | 6,2 | 8,6 | |||
| Широколиственные листопадные леса | 7,6 | 11,4 | |||
| Субтропические леса | 5,3 | 11,3 | |||
| Влажные тропические леса | 10,3 | 37,1 | |||
| Саванна, чапаррель | 6,2 | 9.2 | |||
| Степи, прерии | 2,8 | 3,6 | |||
| Пустыни | 22,7 | 4,2 | |||
| Пашня, обрабатываемые земли | 15,1 | 21,1 . | |||
| Освоенные и окультуренные пастбища | 26,3 | 18,3 | |||
| Воды суши | 2,4 | 0,5 | |||
| Сооружения, дороги, горные выработки | 9,8 | ||||
| Полярные и горные льды | 17,2 | ||||
| Итого для всей суши | 148,9 | ||||
| Океан | 361,1 | ||||
| Всего | 510,0 |
В тех сообществах, где преобладают однолетние растения, травы (тундра, степи, обрабатываемые земли), годовая продукция мало отличается от среднегодовой биомассы. В лесах же продукция составляет лишь от 3,5 до 6,7% от биомассы. В океане продукция заметно превышает среднегодовую биомассу.
Биосфера
Пространство, занимаемое современной биосферой, охватывает приземный слой атмосферы, поверхностные горизонты земной коры континентов и гидросферу Земли. Верхняя граница основного слоя биосферы расположена на высоте нескольких десятков метров над поверхностью растительного покрова на суше или над океаном; нижняя - по горизонту грунтовых вод или максимального проникновения корней растений и роющих животных. В океане она ограничена слоем проникновения солнечных лучей, достаточным для осуществления фотосинтеза (не более 100 м) или глубиной сохранения биологической активности в донных осадках. За этими пределами остается ничтожная часть живых организмов, но находятся огромные массы продуктов их жизнедеятельности - ив атмосфере (газы, пары воды), и в гидросфере (растворенная и взвешенная органика).
В таблице 3.2 сопоставлены некоторые количественные характеристики биосферы и других геосфер Земли. Масса живого вещества биосферы сравнительно мала. Если ее распределить по всей поверхности планеты, то получится слой всего в 1,5 см. Эта «пленка жизни» (выражение В.И.Вернадского), составляя менее 10"6 массы других оболочек Земли, обладает несравненно большим разнообразием, и обновляет свой состав в миллион раз быстрее.
Биота биосферы обусловливает преобладающую часть химических превращений на планете. Отсюда суждение В.И.Вернадского об огромной преобразующей геологической роли живого вещества. На протяжении органической эволюции живые организмы тысячекратно (для разных круговоротов от 103 до 105) пропустили через себя, через свои органы, ткани, клетки, кровь всю атмосферу, весь объем Мирового океана, большую часть массы почв, огромную массу минеральных веществ. И не только «пропустили», но и в соответствии со своими потребностями видоизменили всю земную среду.
Таблица 3.2
Сравнение биосферы с другими геосферами Земли
| Геосферы | Масса, т | Разнообразие состава, V* | Время оборота состава, лет |
| Литосфера (кора выветривания) | 2,5 *1018 | 1,85 | 5*107 |
| Гидросфера | 1,4 *1018 | 0.12 | 2*104 |
| Атмосфера | 5,2 *1015 | 0,38 | 3*104 |
| Биота биосферы** | 2,1 *1012 | 4,50 |
*Индекс разнообразия по Шеннону
** Живое вещество в расчете на сухой вес
Дата добавления: 2016-11-04; просмотров: 3132;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
