ГЛАВА 2. МЕТАБОЛИЗМ. ПРИНЦИПЫ РЕГУЛЯЦИИ ОБМЕНА ВЕЩЕСТВ МИКРОРГАНИЗМОВ
Метаболизмом или обменом веществ называется сумма целенаправленных реакций, протекающих под действием ферментных систем клетки, которые регулируются различными внешними и внутренними факторами, и обеспечивающие обмен веществами и энергией между средой обитания и клеткой.
Вся совокупность химических реакций в клетке (метаболизм) подчиняется принципу биохимического единства – в биохимическом отношении все живые существа на Земле сходны. У них единообразие строительных блоков, единая «энергетическая валюта» (АТФ), универсальный генетический код и в основе своей идентичны главные метаболические пути.
Реакции, приводящие к расщеплению и окислению веществ с получением энергии, называются катаболизмом; пути, приводящие к синтезу основных сложных веществ, называют анаболизмом. Катаболизм и анаболизм – два самостоятельных пути в обмене веществ, хотя отдельные участки их могут быть общими. Такие общие участки, свойственные катаболизму и анаболизму, называются амфиболитическими.
Катаболитические и анаболитические превращения осуществляются последовательно, так как продукт реакции предыдущей стадии является субстратом для последующей.
Энергетический обмен тесно связан с конструктивным (рис. 2.1).
В ходе биологического окисления образуются разнообразные промежуточные продукты (фосфорные эфиры сахаров, пировиноградная, уксусная, щавелевоуксусная, янтарная, a-кетоглутаровая кислоты), из которых вначале синтезируются монополимеры (аминокислоты, азотистые основания, моносахариды), а затем основные макромолекулы клетки. Синтез компонентов клетки идет с затратой энергии, которая образуется при энергетическом обмене. Эта энергия затрачивается также на осуществление активного транспорта веществ, необходимых для анаболизма.
Взаимосвязь конструктивного и энергетического обмена заключается и в том, что процессы биосинтеза, кроме энергии, требуют поступления извне восстановителя в виде водорода, источником которого также служат реакции энергетического обмена.
Скорость течения реакций и в целом обмен веществ клетки зависят от состава питательной среды, условий культивирования микроорганизмов и, главное, от потребности клетки в каждый данный момент в энергии (АТФ) и биосинтетических структурах. Клетка очень экономно высвобождает энергию, и синтезируют веществ ровно столько, сколько необходимо ей в настоящий момент. Этот принцип лежит в основе регуляции и контроля всех стадий метаболических путей в клетке.
Регуляция метаболизма в микробной клетке имеет сложную взаимозависимую систему, которая «включает» и «выключает» определенные ферменты с помощью самых различных факторов: рН среды, концентрации субстратов, некоторых промежуточных и конечных метаболитов и т.д. Изучение путей регуляции определенных продуктов обмена веществ в клетке открывает неограниченные возможности для определения оптимальных условий биосинтеза микроорганизмами целевых продуктов.
гидролитические ферменты
ферменты дальнейших превращений
окислительно-восстановительные
|





 продуктов гидролиза А
продуктов гидролиза А


 ферменты
ферменты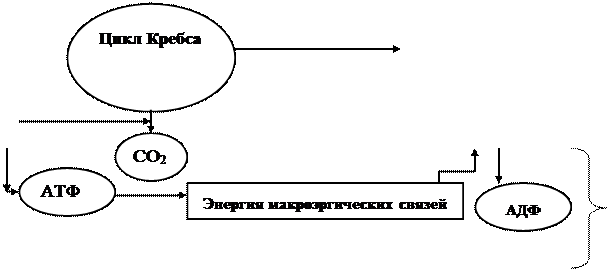
 Б
Б

Рис.2.1. Схема катаболизма и анаболизма микробной клетки
А – конструктивный обмен; Б – энергетический обмен
Для существования жизни важны как регуляция активности отдельных путей метаболизма, так и координация деятельности этих путей.
Каждое из множества веществ создается в клетке в строго необходимых для роста пропорциях в результате ферментативных реакций. Ферменты, синтезирующиеся в клетке постоянно и образование которых не зависит от состава питательной среды называют конститутивными, например, ферменты гликолиза. Другие энзимы, адаптивные или индуцибельные, возникают только в ответ на появление в питательной среде индукторов – субстратов или их структурных аналогов.
Координация химических превращений, обеспечивающая экономность метаболизма, осуществляется у микроорганизмов тремя основными механизмами:
· регуляцией активности ферментов, в том числе путем ретроингибирования;
· регуляцией объема синтеза ферментов (индукция и репрессия биосинтеза ферментов);
· катаболитной репрессией.
В процессе ретроингибирования(ингибирование по принципу обратной связи) активность фермента (аллостерического белка), стоящего в начале многоступенчатого превращения субстрата, тормозится конечным метаболитом, например:
 Аспартат →Карбамил аспартат →Дигидро-оротовая кислота →Оротовая кислота →
Аспартат →Карбамил аспартат →Дигидро-оротовая кислота →Оротовая кислота →

 → Оротидин монофосфат → УМФ → ЦТФ
→ Оротидин монофосфат → УМФ → ЦТФ
Карбамилтрансфераза
 Хоризмат →Антранилат → Индолил глицерофосфат →Триптофан
Хоризмат →Антранилат → Индолил глицерофосфат →Триптофан


 Антранилатсинтетаза
Антранилатсинтетаза
Низкомолекулярные метаболиты передают информацию об уровне своей концентрации и состоянии обмена веществ ключевым ферментам метаболизма. Ключевые ферменты – это регуляторы периодичности образования продукта. С помощью описанного механизма конечные продукты саморегулируют свой биосинтез. Ретроингибирование – способ точного и быстрого регулирования образования продукта. На обмен веществ, аналогичный конечным метаболитам, оказывают эффект их аналоги.
Регуляция объема биосинтеза ферментов (индукция и репрессия) осуществляется на оперонном уровне (Ф.Жакоб и Ж.Моно, 1961) путем изменения количества иРНК, образующихся в процессе транскрипции.
Бактериальная клетка имеет множество генов, каждый из которых несет информацию и контролирует синтез одного белка или соответствующего соединения. Гены подразделяются на структурные гены, гены-регуляторы и гены-операторы. В структурных генах закодирована информация о первичном строении контролируемого ими белка, т.е. о последовательности расположения аминокислот, входящих в состав белка. Гены-регуляторы контролируют синтез белков-репрессоров, подавляющих функцию структурных генов, а гены-операторы выполняют роль посредников между генами регуляторами и структурными генами. (рис. 2.2).
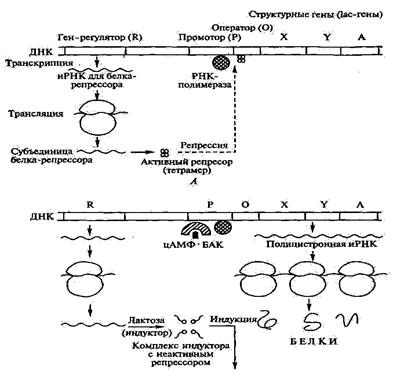
| Опероном называется упорядоченная совокупность структурных генов (со знаками начала и конца) и регуляторных участков. В состав регуляторной зоны оперона входят ген-регулятор, промотор, усилители транскрипции (энхансеры), ослабители транскрипции (сайлансеры) и другие компоненты. Экспрессия регуляторного гена приводит к синтезу белка-репрессора, |
| Рис.2. Структура и механизм индукции и репрессии lac-оперона (по Т.А. Егорова, 2003) А – в отсутствие индуктора; Б – в присутствии индуктора и при дефиците глюкозы |
который в свою очередь способен оккупировать зону первоначального связывания РНК-полимеразы (оператор) тем самым, препятствуя связыванию последней с промоторным участком и началу синтеза иРНК. Конечные продукты метаболических путей могут не только ингибировать активность ферментов первых стадий процесса, но и тормозить биосинтез ферментов последних его этапов, активируя белок репрессор.
Обнаруженный феномен назван репрессией, а ферменты, биосинтез которых стопорится под влиянием низкомолекулярных метаболитов, переводящих репрессорный белок в активную форму, называются репрессибельными. К их числу относятся глутаминсинтетаза, триптофансинтетаза, орнитин-карбамилтрансфераза, уреаза и др. Если концентрация конечного продукта уменьшается до определенного очень низкого уровня, то происходит дерепрессия фермента, т. е. скорость их биосинтеза возрастает до необходимых величин.
В процессе индукции низкомолекулярный метаболит-индуктор (например, лактоза), соединяясь с репрессорным белком (продукт гена-регулятора), инактивирует его и тем самым препятствует взаимодействию белка-репрессора с зоной оператора, что обеспечивает возможность присоединения к промотору РНК-полимеразы и начало синтеза иРНК. Бактериальные клетки продуцируют множество низкомолекулярных эффекторов в ответ на изменение окружающей среды (стресс, голодание, действие фагов и пр.). Каждый из эффекторов, взаимодействуя по аллостерическому механизму с определенными регуляторными белками, моделирует промоторную специфичность РНК-полимеразы, запуская тем самым экспрессию определенного набора генов.
Катаболитная репрессия. Сущность катаболитной репрессии заключается в подавлении биосинтеза ферментов, обеспечивающих метаболизм одного источника углерода другим источником углерода. Ранее считали, что причина такой репрессии состоит в подавлении биосинтеза ферментов обмена одного источника углерода, продуктами катаболизма другого.
Если в питательной среде присутствуют несколько различных источников углерода, клетка микроорганизма вырабатывает ферменты для усвоения лишь одного, наиболее предпочтительного субстрата. Так, например, когда клетки выращивают на смеси глюкозы и лактозы, то в первую очередь утилизируется глюкоза. После полного использования глюкозы происходит экспрессия ферментов метаболизма лактозы (экспрессия структурных генов лактозного оперона). Лактозный оперон (lac-оперон) включает структурные гены трех ферментов: X, Y и А (отвечают за взаимозависимый синтез β-галактозидазы, галактозилпермеазы и ацетилтрансферазы), контролирующих метаболизм лактозы в клетке. Об отсутствие глюкозы в среде сигнализирует цАМФ, синтез которой подавляется в присутствии глюкозы. Уровень цАМФ в клетке является функцией активности аденилатциклазы. цАМФ является необходимым компонентом для связывания РНК-полимеразы с зоной промотора и начала транскрипции генов, ответственных за синтез данных ферментов. В присутствии глюкозы концентрация цАМФ недостаточна для образования комплекса.
Итак, задача регуляторных механизмов заключается в эффективном регулировании и координировании путей метаболизма с целью поддержания необходимой концентрации клеточных компонентов. Кроме того, клетки должны адекватно реагировать на изменения условий окружающей среды посредством включением новых катаболических путей направленных на использование, имеющихся на данный момент питательных субстратов. Регуляция важна для поддержания баланса между энергетическими и синтетическими реакциями в клетке.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ:
1. В чем сущность энергетического обмена?
2. В чем состоит взаимосвязь конструктивного и энергетического обмена?
3. Что такое «фосфорилирование»?
4. Какие ферменты принимают участие в энергетическом обмене аэробов, факультативных анаэробов, облигатных анаэробов?
5. Что подразумевается под «амфиболитическими путями»?
6. Ферменты, и их биохимическая роль.
7. Классификация и номенклатура ферментов.
8. Активные центры ферментов. Субстратная специфичность.
9. Факторы, обеспечивающие ферментативный катализ.
10. Охарактеризуйте состояние равновесия ферментативной реакции?
11. Почему ферменты ускоряют реакции? Что такое энергия активации?
12. От чего зависит скорость ферментативной реакции?
13. Что такое специфичность ферментов?
14. Как называются ферменты, которые выделяются во внешнюю среду?
15. Что такое индуцибельные ферменты?
16. Что такое конститутивные ферменты?
17. Что такое коферменты? Назовите их классы.
18. Как называются ферменты, катализирующие синтетические процессы?
19. Что такое ретроингибирование?
20. Суть теории регуляции синтеза ферментов Ф.Жакоба и Ж.Моно.
21. Объясните механизм индукции синтеза ферментов.
22. Объясните механизм репрессии синтеза ферментов.
23. Что такое катаболитная репрессия?
Дата добавления: 2016-10-26; просмотров: 5223;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. Этические принципы психолога
- I.2.2 ТЕПЛОЁМКОСТЬ ВЕЩЕСТВА
- II. Общие методические принципы в канистерапии
- II. Поведение веществ в магнитном поле.
- III. Механизмы регуляции количества ферментов
- IV. Ликвидация гиповолемии и регуляция водно-солевого обмена
- VI. Биоэнергетические принципы аналитической терапии

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
