Изменения структурной организации хромосом. Хромосомные мутации
Несмотря на эволюционно отработанный механизм, позволяющий сохранять постоянной физико-химическую и морфологическую организацию хромосом в ряду клеточных поколений, под влиянием различных воздействий эта организация может изменяться. В основе изменения структуры хромосомы, как правило, лежит первоначальное нарушение ее целостности — разрывы, которые сопровождаются различными перестройками, называемыми хромосомными мутациями или аберрациями.
Разрывы хромосом происходят закономерно в ходе кроссинговера, когда они сопровождаются обменом соответствующими участками между гомологами (см. разд. 3.6.2.3). Нарушение кроссинговера, при котором хромосомы обмениваются неравноценным генетическим материалом, приводит к появлению новых групп сцепления, где отдельные участки выпадают — делении — или удваиваются — дупликации (рис. 3.57). При таких перестройках изменяется число генов в группе сцепления.
Разрывы хромосом могут возникать также под влиянием различных мутагенных факторов, главным образом физических (ионизирующего и других видов излучения), некоторых химических соединений, вирусов.
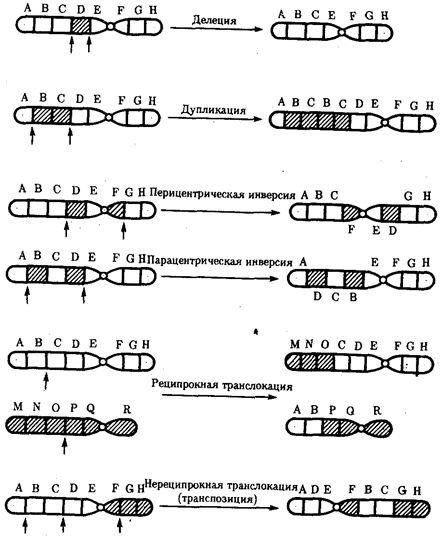
Рис. 3.57. Виды хромосомных перестроек
Нарушение целостности хромосомы может сопровождаться поворотом ее участка, находящегося между двумя разрывами, на 180° — инверсия. В зависимости от того, включает ли данный участок область центромеры или нет, различают перицентрические и парацентрические инверсии (рис. 3.57).
Фрагмент хромосомы, отделившийся от нее при разрыве, может быть утрачен клеткой при очередном митозе, если он не имеет центромеры. Чаще такой фрагмент прикрепляется к одной из хромосом — транслокация. Нередко две поврежденные негомологичные хромосомы взаимно обмениваются оторвавшимися участками —ре-ципрокная транслокация (рис. 3.57). Возможно присоединение фрагмента к своей же хромосоме, но в новом месте — транспозиция (рис. 3.57). Таким образом, различные виды инверсий и транслокаций характеризуются изменением локализации генов.
Хромосомные перестройки, как правило, проявляются в изменении морфологии хромосом, что можно наблюдать в световой микроскоп. Метацентрические хромосомы превращаются в субметацентрические и акроцентрические и наоборот (рис. 3.58), появляются кольцевые и полицентрические хромосомы (рис. 3.59). Особую категорию хромосомных мутаций представляют аберрации, связанные с центрическим слиянием или разделением хромосом, когда две негомологичные структуры объединяются в одну — робертсоновская транслокация, или одна хромосома образует две самостоятельные хромосомы (рис. 3.60). При таких мутациях не только появляются хромосомы с новой морфологией, но и изменяется их количество в кариотипе.
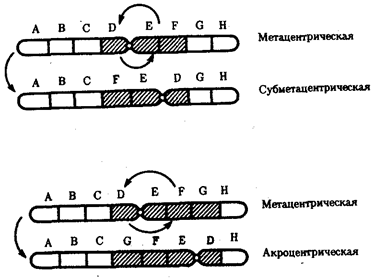
Рис. 3.58. Изменение формы хромосом
в результате перицентрических инверсий
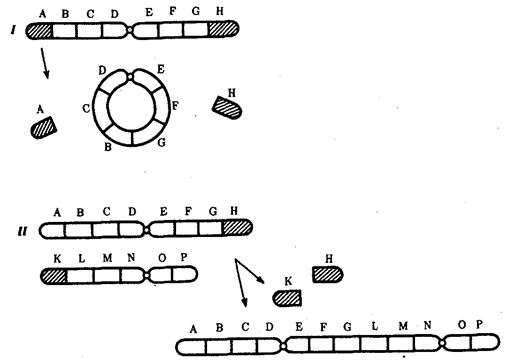
Рис. 3.59. Образование кольцевых (I) и полицентрических (II) хромосом
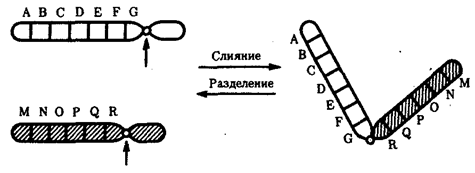
Рис. 3.60. Хромосомные перестройки, связанные с центрическим слиянием
или разделением хромосом являются причиной изменения числа хромосом
в кариотипе
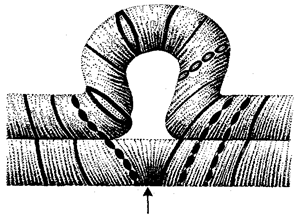
Рис. 3.61. Петля, образующаяся при конъюгации гомологичных хромосом, которые несут неравноценный наследственный материал в соответствующих участках в результате хромосомной перестройки
Описанные структурные изменения хромосом, как правило, сопровождаются изменением генетической программы, получаемой клетками нового поколения после деления материнской клетки, так как изменяется количественное соотношение генов (при делениях и дупликациях), меняется характер их функционирования в связи с изменением взаимного расположения в хромосоме (при инверсии и транспозиции) или с переходом в другую группу сцепления (при транслокации). Чаще всего такие структурные изменения хромосом отрицательно сказываются на жизнеспособности отдельных соматических клеток организма, но особенно серьезные последствия имеют хромосомные перестройки, происходящие в предшественниках гамет.
Изменения структуры хромосом в предшественниках гамет сопровождаются нарушением процесса конъюгации гомологов в мейозе и их последующего расхождения. Так, делении или дупликации участка одной из хромосом сопровождаются при конъюгации образованием петли гомологом, имеющим избыточный материал (рис. 3.61). Реципрокная транслокация между двумя негомологичными хромосомами приводит к образованию при конъюгации не бивалента, а квадривалента, в котором хромосомы образуют фигуру креста благодаря притягиванию гомологичных участков, расположенных в разных хромосомах (рис. 3.62). Участие в реципрокных транслокациях большего числа хромосом с образованием поливалента сопровождается формированием еще более сложных структур при конъюгации (рис. 3.63).
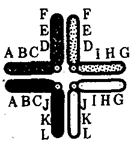 Рис. 3.62. Образование при конъюгации квадривалента из двух пар хромосом, несущих реципрокную транслокацию
Рис. 3.62. Образование при конъюгации квадривалента из двух пар хромосом, несущих реципрокную транслокацию
| 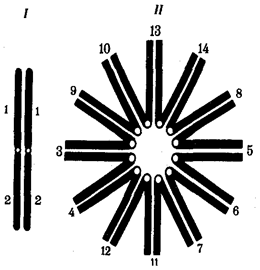 Рис. 3.63. Образование при конъюгации поливалента шестью парами хромосом, участвующих
в реципрокных транслокациях:
I — конъюгация между парой хромосом, не несущих транслокацию;
II — поливалент, образуемый шестью парами хромосом, участвующих
в транслокации
Рис. 3.63. Образование при конъюгации поливалента шестью парами хромосом, участвующих
в реципрокных транслокациях:
I — конъюгация между парой хромосом, не несущих транслокацию;
II — поливалент, образуемый шестью парами хромосом, участвующих
в транслокации
|
В случае инверсии бивалент, возникающий в профазе I мейоза, образует петлю, включающую взаимно инвертированный участок (рис. 3.64).
Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению новых хромосомных перестроек. В результате гаметы, получая неполноценный наследственный материал, не способны обеспечить формирование нормального организма нового поколения. Причиной этой является нарушение соотношения генов, входящих в состав отдельных хромосом, и их взаимного расположения.
Однако, несмотря на неблагоприятные, как правило, последствия хромосомных мутаций, иногда они оказываются совместимыми с жизнью клетки и организма и обеспечивают возможность эволюции структуры хромосом, лежащей в основе биологической эволюции. Так, небольшие по размеру делении могут сохраняться в гетерозиготном состоянии в ряду поколений. Менее вредными, чем делении, являются дупликации, хотя большой объем материала в увеличенной дозе (более 10% генома) приводит к гибели организма.
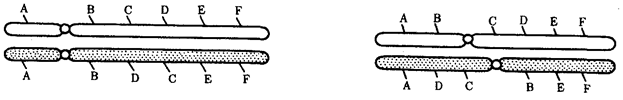
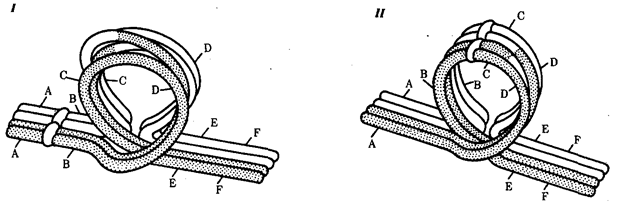
Рис. 3.64. Конъюгация хромосом при инверсиях:
I — парацентрическая инверсия в одном из гомологов, II — перидентрическая инверсия в одном из гомологов
Нередко жизнеспособными оказываются робертсоновские транслокации, часто не связанные с изменением объема наследственного материала. Этим можно объяснить варьирование числа хромосом в клетках организмов близкородственных видов. Например, у разных видов дрозофилы количество хромосом в гаплоидном наборе колеблется от 3 до 6, что объясняется процессами слияния и разделения хромосом. Возможно, существенным моментом в появлении вида Homo sapiens были структурные изменения хромосом у его обезьяноподобного предка. Установлено, что два плеча крупной второй хромосомы человека соответствуют двум разным хромосомам современных человекообразных обезьян (12-й и 13-й —шимпанзе, 13-й и-14-й —гориллы и орангутана). Вероятно, эта человеческая хромосома образовалась в результате центрического слияния по типу робертсоновской транслокации двух обезьяньих хромосом.
К существенному варьированию морфологии хромосом, лежащему в основе их эволюции, приводят транслокации, транспозиции и инверсии. Анализ хромосом человека показал, что его 4, 5, 12 и 17-я хромосомы отличаются от соответствующих хромосом шимпанзе перицентрическими инверсиями.
Таким образом, изменения хромосомной организации, чаще всего оказывающие неблагоприятное воздействие на жизнеспособность клетки и организма, с определенной вероятностью могут быть перспективными, наследоваться в ряду поколений клеток и организмов и создавать предпосылки для эволюции хромосомной организации наследственного материала.
Дата добавления: 2016-08-23; просмотров: 2524;
Поиск по сайту
Узнать еще
- C учетом изменения статической работы балки
- Cущность организации и ее основные признаки
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- II. Оппозиционные организации и их роль в подготовке революции
- III. Уровни организации живого
- III. Формирование личности посредством профориентации и организации досуговой деятельности.
- PR – подразделение в организации.
- VII.2. Закономерности изменения свойств нефтей и газов на месторождениях

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
