Взаимоотношения рыб с другими животными и растениями.
БИОТИЧЕСКИЕ ВЗАИМООТНОШЕНИЯ У РЫБ
Биотические взаимоотношения у рыб многообразны. Здесь прежде всего следует выделить внутривидовые, межвидовые взаимоотношения, а также взаимоотношения рыб с другими водными животными и растениями.
Внутривидовые взаимоотношения у рыб.Внутривидовые связи у рыб проявляются в образовании стад, элементарных популяций, стай, скоплений, колоний, а также в пищевых взаимоотношениях.
Популяция, или стадо — это одновидовая разновозрастная самовоспроизводящаяся группировка рыб, приуроченная к определенному месту обитания и характеризующаяся определенными морфобиологическими показателями — размерно-возрастным составом, темпом роста, сроками нереста и т.д.
Например, вобла в Каспийском море образует северокаспийское, куринское и туркменское стада. Морской судак в Южном Каспии также представлен несколькими стадами. Тихоокеанская сельдь распадается на большое количество локальных стад, промысловое значение из которых имеют декастринское, стадо сельди залива Петра Великого, сахалино-хоккайдское, гижигинское и корфо-карагинское (рис. 1).
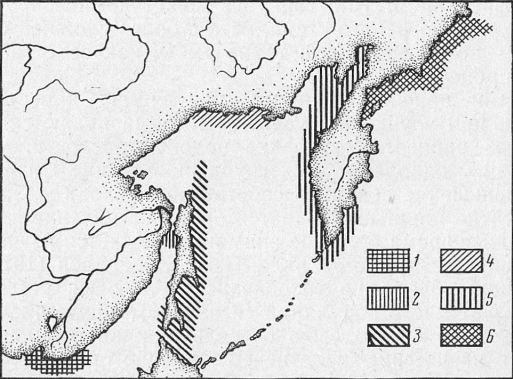
Рис. 1. Ареалы стад тихоокеанской сельди (Правоторова):
1—сельдь залива Петра Великого; 2 — декастринское; 3 — сахалино-хоккайдское; 4 — охотское; 5 — гижигинское; 6 — корфо-карагинское.
Жизнь популяции состоит в непрерывной смене поколений, их росте, созревании, воспроизводстве потомства и смерти. Каждая популяция, как и вид в целом, представляет собой саморегулирующуюся систему, находящуюся в постоянном взаимодействии с абиотическими и биотическими факторами, которые влияют на все этапы жизненного цикла рыб и сказываются на численности популяции. Популяция является основной структурной единицей вида, на которой резко ощущается воздействие промысла, хищников, паразитов и болезней, обеспеченность пищей, а также изменение абиотических условий.
Колебания численности популяции в значительной степени зависят от выживания икринок и личинок на ранних стадиях развития, интенсивности промысла и обеспеченности рыб пищей.
В настоящее время интенсивный промысел с применением высокоэффективных орудий лова и поисковой техники, позволяющей обнаруживать рыбу в толще воды, в ряде случаев является важнейшим фактором, определяющим изменение численности и структуры популяций промысловых рыб. В последние годы весьма нарушилась структура и сократилась численность популяций многих промысловых рыб — атлантической трески, пикши, атлантической сельди, морского окуня, большинства камбал, лососей и др.
Промысел, воздействуя на определенную часть популяции, может изменять ее структуру (возрастной состав, соотношение полов) , значительно «омолаживая» ее.
Рыбы с коротким жизненным циклом, ранней половой зрелостью, т. е. с большой воспроизводительной способностью и относительно простой структурой популяции, могут переносить большее промысловое изъятие, чем рыбы с длинным жизненным циклом и поздним созреванием.
Так, горбуша может компенсировать изъятие до 60% половозрелого стада, кета с более сложной структурой популяции — не более 50%, а такие рыбы, как осетровые, — всего несколько процентов.
Основными приспособлениями популяции к регулированию численности при улучшении условий откорма являются ускорение роста, более раннее наступление половой зрелости, увеличение плодовитости у одноразмерных групп, повышение жизнестойкости молоди, уменьшение доедания собственной молоди у хищных рыб. При недостатке пищи наблюдается обратное явление.
В последнее время большое внимание уделяется изучению внутрипопуляционной структуры вида. С. М. Коновалов (1979), изучая популяционную биологию тихоокеанских лососей, установил, что нерка образует много популяций (изолятов), общее число которых составляет около 130. В свою очередь каждая популяция может распадаться на весенне- и летненерестующие расы, приспособленные к определенным условиям инкубации икры и разновременному выклеву личинок. Каждая сезонная раса состоит из субпопуляций (субизолятов), количество которых в значительной мере зависит от числа и особенностей нерестовых речек и озерных нерестилищ.
В бассейне оз. Азабачьего (Камчатка), например, выделено до 40 субпопуляций нерки. Субпопуляции различаются по генетической, размерно-весовой, возрастной, половой и трофической структурам и характеризуются самовоспроизводимостью.
Знание субпопуляционной структуры используемого промыслом вида имеет большое значение для осуществления рациональной его эксплуатации.
Элементарные популяции, впервые выделенные у рыб В.Д. Лебедевым (1967), представляют собой группировки, состоящие в основном из рыб одного возраста, близких по физиологическому состоянию (упитанность, степень зрелости гонад, количество гемоглобина в крови, зараженность паразитами) и сохраняющихся пожизненно. Элементарными их называют потому, что они не распадаются ни на какие внутривидовые биологические группировки.
Элементарные популяции возникают в местах рождения, потому что отдельные группировки в результате разнокачественности икры проходят одни и те же этапы развития в разное время и с разной скоростью.
Элементарные популяции обнаружены у самых различных видов рыб — азовской хамсы, северокаспийской и аральской воблы, тюльки, красноперки, морского окуня, трески Баренцева моря и др.
Структура элементарных популяций изменчива. Они могут образовывать стаи различной величины или сливаться в одно большое скопление, простирающееся на 20—30 миль и более, или какое-то время находиться в разреженном состоянии, а затем вновь образовывать стаи. Поэтому эти группировки нельзя назвать ни стаями, ни скоплениями, так как они существуют и тогда, когда находятся в разреженном состоянии.
В элементарные популяции объединяются рыбы с одинаковым сезонным ритмом жизнедеятельности. Они устойчивы и могут быть прослежены в течение длительного времени.
Элементарная популяция и стая — группировки разного порядка. Если элементарная популяция представляет собой единицу популяционной структуры вида и является частью стада, то стая связана с поведением рыб.
Большинству видов рыб свойствен одиночный образ жизни (щука, сом, скаты, гигантская акула, луна-рыба и др.). Из всего многообразия рыб (более 20 тыс. видов) лишь около 4 тыс. видов являются стайными.
Стая (косяк) — это группировка близких по возрастному и биологическому состоянию рыб, объединяющихся на более или менее длительный период. Стайными являются в основном пелагические рыбы. Наиболее отчетливо стайность выражена у анчоусовых, сельдевых, ставридовых, скумбриевых рыб. Большие стаи образуют полупроходные рыбы — вобла, лещ, судак и др.
Свойство стайности сложилось у рыб в процессе длительного развития как приспособление, обеспечивающее лучшую выживаемость, так как стайный образ жизни способствует поиску пищи, защите от хищников, нахождению миграционных путей. Поведение стаи напоминает единый организм, а ее устойчивость в первую очередь обусловлена зрительными контактами, чему способствуют стайная окраска у некоторых рыб (у пикши черное пятно над грудными плавниками), а также издаваемые стайными рыбами звуки и образуемые ими электрические поля.
Форма, величина, плотность и структура стай даже у одного вида рыб бывают различными, но вместе с тем при сходных условиях стаи разных видов рыб могут иметь много общего. Стаи многих видов рыб (ставрида, сардина, сельдь, пеламида) многослойные, при быстром движении имеют форму клина, а при откорме —округлую.
Стаи каспийской кефали однослойные, плоские, а форма их может быть округлой, подковообразной или лентообразной. Общая протяженность цепи косяков кефали может достигать 100 км.
Количество рыб в стае различно — от нескольких особей до огромного количества. Так, площадь стай зимующей хамсы в Черном море в 1952 г. изменялась от 3,4 до 444,4 тыс. м2 при средней высоте 14,0—23,5 м. Объем этих косяков достигал 8,7 млн. м3.
Структура стай (взаимное расположение особей в пространстве) также различна и связана с действием рыб в данный момент. Различают следующие типы структуры стай рыб: ходовая (все рыбы движутся в одном направлении), кругового обзора (рыбы малоподвижны и ориентированы в разные стороны), оборонительная (стая стремится ускользнуть от хищника), питающаяся.
Защитное значение стаи пелагических рыб от дневных хищников при высокой освещенности заключается в том, что рыбы, находящиеся в стае, менее доступны для них. Стая рыб раньше замечает опасность, а между тем отдельные движущиеся рыбы в стае трудноразличимы, и невозможно остановить взгляд на одной рыбе.
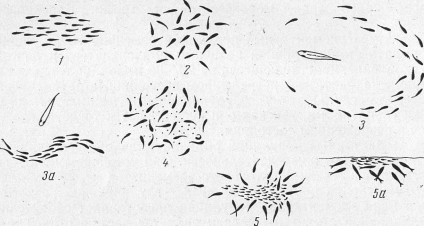
Рис. 2. Основные типы структуры стаи рыб (Радаков):
1 - ходовая; 2- кругового обзора; 3 и За - оборонительная; 4 - при питании рыб-планктофагов; 5 - при питании пелагических хищников (5а - вид сбоку).
Бросок вслепую бывает в основном безуспешным, так как стая разделяется на две части и перед хищником образуется пустота. Поэтому хищник стремится отбить от, стаи отдельных рыб, а затем уже пытается схватить их. Иногда голодные хищники могут подолгу кружить вокруг стаи, не нападая на рыб.
Реакция на опасность у стайных рыб различна. Стая анчоуса при нападении хищника превращается в плотный шар, а стая скумбрии рассеивается и опускается на глубину, что дезориентирует хищника.
Пелагические стайные рыбы, ориентирующиеся в основном при помощи органов зрения, днем держатся стаями, питаясь и спасаясь от хищников. Ночью стаи рассредоточиваются, так как, находясь в стае, рыбы более доступны для ночных хищников, чем одиночные особи или объединившиеся в мелкие косячки.
Изучение поведения рыб в зоне действия орудий лова имеет важное значение для промышленного рыболовства. Поймать косячную рыбу движущимися орудиями лова значительно труднее, чем рассредоточенную, так как стая на большем расстоянии, чем одиночные рыбы, замечает орудие лова и стремится избежать его.
Большие уловы стайных рыб обычно получают в том случае, если орудие лова охватывает всю стаю.
Стайное поведение некоторых хищников облегчает им поимку добычи, так как они могут оттеснить ее от обычных укрытий, окружить, лишив возможности спастись бегством.
Приспособительное значение имеет стая и при миграциях. Как уже указывалось, биоэлектрические поля отдельных рыб в стае суммируются, и образуется единое биоэлектрическое поле стаи, благодаря которому стая ориентируется в пространстве и находит необходимые для данного периода места обитания.
Знание закономерностей образования стай, их распада, передвижения, реакции на орудия лова имеет важное значение для организации эффективного промысла.
Скопление — это огромная масса рыбы, представляющая собой ряд стай, которые могут сливаться или быть обособленными.
Состав рыб в скоплениях часто бывает разнокачественным.
Скопления бывают нерестовые, нагульные, миграционные и зимовальные. Огромные зимовальные скопления образует, например, хамса в Черном море. Такие скопления хамсы обычно малоподвижны. При переходе из одного района в другой скопление распадается на отдельные косяки, которые после прекращения движения сливаются снова.
Нерестовые скопления образуются на местах размножения и состоят в основном из половозрелых особей, например скопления сельди, трески или мойвы у берегов Норвегии. Нагульные скопления образуются на местах откорма рыб. Они могут состоять из рыб разных видов и разного возраста. Примером могут служить нагульные скопления атлантической сельди в Норвежском море.
Миграционные скопления возникают на путях движения рыб на нерест, нагул или зимовку, например скопления азовской хамсы, двигающейся из Азовского моря в Черное на зимовку.
Колонии — это временные защитные группировки рыб, обычно состоящие из особей одного пола. Они образуются на местах размножения для защиты кладок икры от врагов. Колонии известны у косатки-скрипуна, панцирных американских сомов (Loricariidae) и др.
Внутривидовой паразитизм, являющийся одним из проявлений многогранных взаимоотношений у рыб, наблюдается у глубоководных удильщиков, крошечные самцы которых прирастают к телу самки и питаются через ее кровеносную систему (рис. 3).
Важное значение в жизни рыб имеют внутривидовые пищевые взаимоотношения . Путем ряда приспособлений достигаются ослабление напряженности внутривидовых пищевых отношений и обеспечение популяции пищей. В частности, ослабление внутривидовых противоречий из-за пищи связано с такими приспособлениями, как наличие у ряда видов нескольких генераций молоди в течение года, образующихся в результате порционного нереста, из-за расхождения в составе пищи на разных этапах развития у молоди и взрослых рыб, у самцов и самок (см. «Питание»).

Рис. 3 Внутривидовой паразитизм.
Кроме того, обычно в условиях плохой обеспеченности пищей наблюдается расширение спектра питания рыб, а в условиях хорошей обеспеченности — сужение.
Приспособлением к условиям откорма взрослых рыб является характер их роста. У многих рыб при ухудшении условий питания наблюдается замедление темпа роста, что ведет к более позднему созреванию, а следовательно, сокращению численности стада в соответствии с кормовой базой. Ускорение роста и созревания рыб в молодом возрасте происходит при улучшенных условиях откорма.
Более рациональное использование кормовых ресурсов обеспечивается также и тем, что у некоторых видов наблюдается большая гибель самцов после нереста (лососи, бычки), так как на восстановление жизненных функций одного старого самца затрачивалось бы больше корма, чем на рост двух молодых.
Приспособлением к сохранению популяции при неблагоприятных условиях питания является также образование карликовых форм, для которых характерно сильное замедление роста и созревание при небольших размерах (карликовые самцы лососей, мелкий карась в некоторых водоемах и др.).
У некоторых хищных рыб (треска, навага, корюшка, щука, речной окунь) при недостатке корма сокращение численности достигается переходом на питание собственной молодью (каннибализм).
Щуке, речному окуню и другим рыбам каннибализм позволяет жить даже в таких водоемах, где нет другой рыбы.
Межвидовые взаимоотношения у рыб.Межвидовые взаимоотношения у рыб довольно разнообразны и проявляются в форме пищевой конкуренции, хищника и жертвы, мирного сожительства, паразитизма и др.
Межвидовые связи рыб лучше всего рассматривать на примере фаунистических комплексов. Фаунистический комплекс — это группа видов, связанных общностью своего географического происхождения, т. е. развитием в одной географической зоне, к абиотическим и биотическим условиям которой они приспособились.
Основные фаунистические комплексы пресноводной и солоноватоводной ихтиофауны СССР (Никольский, 1953):
Бореальный предгорный (хариус, речной гольян и др.)
Бореальный равнинный (щука, карась, окунь, ерш, плотва и др.)
Арктический пресноводный (сиги, нельма, лосось, налим и др.)
Арктический морской (камбалы, рогатка, колюшки и др.)
Понтокаспийский пресноводный (лещ, красноперка, жерех и др.)
Понтокаспийскйй морской (сельдь, бычки, морской судак и др.)
Третичный равнинный пресноводный (сазан, вьюн и др.)
Переднеазиатский (храмуля, быстрянки, гольцы и др.)
Нагорноазиатский (маринка, гольцы, османы и др.)
Туркестанский равнинный (лжелопатоносы, щуковидный жерех, остролучка, гольцы и др.)
Индоафриканский (змееголов, косатки и др.)
Китайский равнинный (амур, верхогляд, колючий горчак, востробрюшка)
Межвидовые взаимоотношения у рыб одного фаунистического комплекса прежде всего характеризуются ослаблением пищевой конкуренции (особенно у взрослых рыб) путем расхождения спектров питания и мест кормления. Конкуренция наблюдается в основном из-за второстепенных кормовых объектов. Так, например, в р. Или балхашская маринка питается растительной пищей, а илийская — хищник. Молодь илийской маринки кормится выше по течению, чем балхашской.
Однако ихтиофауна отдельных водоемов, в том числе и рек, обычно слагается из разных фаунистических комплексов, которые вступают в определенные взаимоотношения. Но и в этом случае наблюдается приспособление к ослаблению пищевой конкуренции.
В реке, например, снижение пищевой конкуренции достигается тем, что разные фаунистическне комплексы занимают в основном разные участки реки. Так, в Амударье нагорно-азиатский комплекс занимает верховье, переднеазнатский и индоафриканский — предгорье, туркестанский — равнинное течение, а понтокаспийский — нижнее течение.
Обостренные пищевые отношения между различными фаунистическими комплексами возникают в местах их соприкосновения и существуют из-за основных кормовых объектов. Так, основными фаунистическими комплексами в Амударье являются туркестанский и понтокаспийский. Аральский жерех конкурирует в питании со щуковидным жерехом из-за чехони и остролучки, сом — с большим лжелопатоносом из-за молоди усача и гольцов, а гольцы — с молодью усача из-за личинок хирономид (рис. 4).
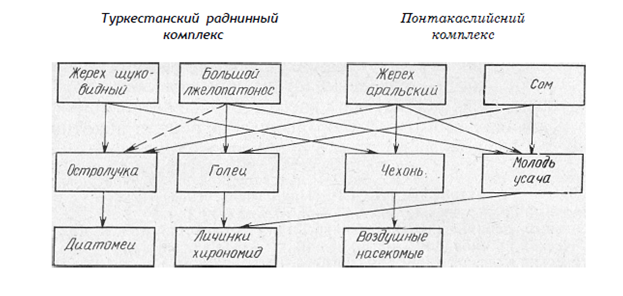
Рис. 4. Пищевые взаимоотношения рыб различных фаунистических комплексов
(по Никольскому).
Характер размножения видов, относящихся к одному фаунистическому комплексу, их плодовитость, наличие гнезд также являются приспособлением к условиям определенной географической зоны и к хищникам. Например, рыбы понтокаспийского фаунистического комплекса мечут икру на растительность (лещ, красноперка, линь и др.), а многие рыбы, входящие в китайский фаунистический комплекс, откладывают икру в толщу воды и в раковины моллюсков, что связано с определенным водным режимом рек этой географической зоны. Так, в Амуре наблюдается два паводка (весенний и летний), и колебания уровней достигают значительной величины. Пелагическая икра проходит свое развитие, сплывая с током воды, что обеспечивает ей лучшую выживаемость по сравнению с икрой, отложенной на растительность.
Взаимоотношения хищника и жертвы привели к выработке у хищников разнообразных приспособлений для добывания пищи — сильных зубов, способности к быстрому передвижению, хорошему обонянию и зрению и т. д., а у потенциальной жертвы появились шипы, колючки, часто снабженные ядовитыми железами. У иглобрюха, например, имеется особый мешок, представляющий собой вырост желудка, который при опасности раздувается, превращая рыбу в своеобразный шар и делая ее недоступной для хищника.
Сложность взаимоотношений хищника и жертвы возрастает потому, что, истребляя разные виды, хищники влияют на межвидовые отношения. Например, в Веселовское водохранилище был вселен судак, который стал питаться в основном мелкими рыбами, конкурирующими в питании с лещом и сазаном. И если до вселения судака на долю леща и сазана приходилось всего 18% улова, а на мелочь — 36%, то после акклиматизации судака соотношение уловов изменилось: более 70% улова составили лещ и сазан, 14% —судак, а мелочь — всего около 3%.
Формы сожительства у рыб различны, это мирное сожительство (комменсализм, симбиоз) и паразитизм.
Примером комменсализма(нахлебничества) могут служить взаимоотношения акул с рыбой прилипалой и рыбой-лоцманом. Используя преобразованный в присоску передний спинной плавник для прикрепления к телу акулы, прилипала следует вместе с ней и отделяется только для того, чтобы съесть остатки пищи.
Рыбы-лоцманы плывут рядом с акулой и при приближении к добыче бросаются вперед, как бы указывая направление к ней, а затем также поедают пищевые остатки.
Симбиоз (сожительство полезное для обоих видов) наблюдается, например, у рыб-чистильщиков с их «клиентами» (рис. 6).
Кожа и жабры рыб, особенно в тропической области, часто поражаются эктопаразитами, грибковыми и бактериальными заболеваниями. Избавиться от них помогают рыбы-санитары, или чистильщики. Известно около 25 видов таких рыб; к ним относятся в основном мелкие окунеобразные рыбы коралловых рифов: губановые, рыбы-бабочки и др. Эти рыбы ярко окрашены и хорошо заметны. Они внимательно осматривают тело «клиента», забираются в рот и жаберную полость, выщипывая паразитов, обкусывая кусочки омертвевшей ткани, снимая грибковый налет. Рыба «клиент» никогда не поедает «чистильщика». Такая санитарная обработка рыб производится в определенных местах, где нередко скапливаются рыбы, нуждающиеся в такой помощи. Один чистильщик за 1 ч может обработать до 50 «клиентов». Исчезновение рыб-чистильщиков приводит к возрастанию степени поражения грибковыми заболеваниями обитающих здесь рыб (Оммани, 1975).
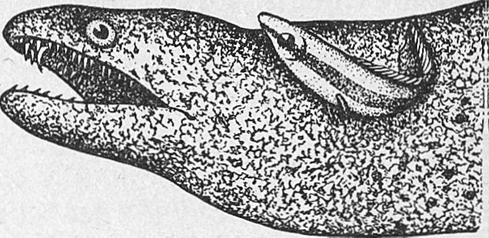
Рис. 5. Радужный губан исцеляет хищную мурену (по Оммани).
Межвидовой паразитизм у рыб встречается сравнительно редко. Так, например, паразитируют на рыбах миксины и миноги, но наиболее яркими паразитами являются маленькие сомики (длиной до 9 см) из семейства ваиделлиевых (Trichomycteridae), обитающие вводоемах Южной Америки, прогрызающие кожу крупных рыб и питающиеся их кровью. Сомик-стегофил паразитирует в жаберной полости крупных сомов и других рыб, где он удерживается при помощи присоски. Сомик-ванделлия паразитирует в мочеполовых протоках крупных рыб, удерживаясь там при помощи шипов жаберной крышки, а также может проникать в мочеполовую систему купающихся людей.
Взаимоотношения рыб с другими животными и растениями.
У рыб существуют тесные связи не только между собой, но и с другими водными животными, как беспозвоночными, так и позвоночными, а также с растениями, бактериями и вирусами.
Многие заболевания рыб имеют бактериальную или вирусную природу. Вирусными заболеваниями являются краснуха и инфекционная водянка у карпов, бактериальными — фурункулез и др. Известны случаи массовой гибели карпа в прудовых хозяйствах и сазана в ряде водоемов (оз. Балхаш и др.) от краснухи, судака в Южном Каспии от фурункулеза.
Некоторые рыбы, в частности белый толстолобик, наряду с фитопланктоном используют в пищу бактерии и в виде бактериальной пленки. Бактерии служат пищей для беспозвоночных.
Светящиеся органы некоторых рыб включают в себя особые бактерии, которые светятся при контакте с кислородом.
Водоросли и высшие растения, выделяя кислород и поглощая углекислый газ, создают благоприятные условия для жизни рыб. Многие рыбы используют растительность как субстрат при откладывании икры (лещ, сазан, вобла, тихоокеанская сельдь и др.). Есть рыбы, которые сооружают гнезда из кусочков растительности (колюшка трехиглая). В зоне макрофитов обитают многие кормовые организмы, а также некоторые рыбы, использующие их для укрытия. Ряд рыб используют плавающие водоросли для откладывания икры (сайра).
Водоросли и высшие растения являются объектами питания растительноядных рыб. Фитопланктоном питаются белый толстолобик, перуанский анчоус, перифитоном, который состоит в основном из нитчатых водорослей, подуст, макрофитами — белый амур, красноперка. Некоторые растения (пузырчатка) питаются личинками рыб.
Из низших растений некоторые грибы вызывают у рыб серьезные заболевания, например бранхиомикоз, или жаберную гниль, и сапролегнию.
Бурное развитие одноклеточных водорослей — динофлагеллят, известное под названием «красное цветение», возникает в некоторых районах Мирового океана в жаркое время и в результате выделяемых ими токсинов может привести к массовой гибели рыб.
Чрезмерное развитие фитопланктона и макрофитов ухудшает кислородный режим водоемов, нарушает равномерное прогревание воды, мешает отлову рыбы.
Молодь почти всех рыб на ранних этапах развития питается простейшими (Protozoa), в основном инфузориями. Однако среди простейших есть немало паразитов, наносящих существенный ущерб рыбному хозяйству.
Кишечнополостные (Coelenterata) лишь в небольшой степени используются рыбами для питания. Только кораллы поедаются некоторыми рыбами и являются убежищем для них. Известно немало примеров сожительства кишечнополостных с рыбами. Так, молодь трески, пикши и других рыб укрывается от хищников под куполом крупной арктической медузы Cyanea capillata и объедает остатки пищи с ее щупалец. Среди щупалец гигантских морских анемон живут рыбки рода Amphiprion.
Некоторые кишечнополостные (гидра, медузы, гребневики) являются хищниками и уничтожают личинок и молодь рыб. Многие из кишечнополостных являются серьезными конкурентами в питании рыб, уничтожая огромное количество зоопланктона, например калянуса — основного компонента питания сельди в Баренцевом море. В Азовском море в связи с повышением солености в последние годы наблюдается массовое развитие медуз, активно выедающих планктон и являющихся большими конкурентом промысловых рыб. Среди кишечнополостных есть и паразитические формы, например Polypodium, поражающий гонады осетровых рыб.
Черви (Vermes) круглые, малощетинковые и многощетинковые имеют важное значение в пище рыб.
Из малощетинковых червей энхитреусом (Enchytraeus albidus) кормят молодь некоторых рыб на рыбоводных заводах. Многощетинковый червь нереис (Nereis succinea), которым в Азовском море питаются многие рыбы, акклиматизированный в Каспийском море, также стал важным кормовым объектом для осетровых, бычков, воблы и др.
В то же время многие черви и особенно ленточные, а также моногенетические и дигенетические сосальщики, скребни и круглые черви являются паразитами, и почти все рыбы в той или иной степени заражены ими.
Моллюски (Mollusca) играют важную роль в питании многих видов рыб — плотвы, воблы, русского осетра, бычков, камбал и др. В мантийную полость двустворчатого моллюска Anodonta горчаки откладывают икру.
В то же время головоногие моллюски, кальмары и каракатицы поедают рыб и в том числе промысловых. Личинки двустворчатых моллюсков Unio u Anodonta глохидин паразитируют на жабрах и плавниках рыб.
Ракообразные (Crustacea) имеют наибольшее значение в питании рыб. Преимущественно ракообразными питается подавляющее большинство мелких пелагических рыб — сельдевых, анчоусовых, скумбриевых и др. Океаническая сельдь, например, питается в основном калянусом (Calanus finmarchicus) и черноглазками (Euphausiacea). В молодом возрасте почти все рыбы, в том числе и хищники потребляют ракообразных.
Наряду с огромным кормовым значением ракообразных для рыб некоторые из них приносят существенный вред рыбному хозяйству. К ним относятся представители отрядов веслоногих (Соpepoda), жаброхвостых (Branchiura) и листоногих (Phyllopoda) ракообразных.
Из веслоногих циклопы, например, нападают на личинок рыб и повреждают их. В некоторые годы на оз. Байкал сильное развитие циклопов в планктоне влечет за собой уменьшение численности эпишуры — основного кормового объекта байкальского омуля.
Ergasilus sieboldi паразитирует на жабрах рыб и вызывает в прудовых хозяйствах массовую гибель рыб. Представитель жаброхвостых карпоед Argulus foliaceus также является одним из серьезных паразитов рыб. Он присасывается к коже рыбы, ослабляет ее, что иногда приводит к массовой гибели молоди. Существенный вред молоди рыб наносят листоногие раки (щитни и др.), являющиеся пищевыми конкурентами и нападающие на мальков рыб. Кроме того, взрыхляя ил, они сильно взмучивают воду, что приводит к загрязнению жабр и массовой гибели мальков. Некоторые ракообразные являются промежуточными хозяевами ремнеца, широкого лентеца и др., выполняя роль переносчиков глистных инвазий (веслоногие рачки).
Насекомые (Insecta) и их личинки, особенно хирономид, ручейников, стрекоз и поденок, являются важными пищевыми объектами пресноводных и солоноватоводных рыб. Немалую роль,в питании некоторых рыб имеют воздушные насекомые. Так, в питании форели в конце лета и осенью упавшие в воду воздушные насекомые составляют около 93%. Некоторые насекомые (водяные жуки и клопы) являются конкурентами рыб в питании, а многие из них, например различные плавунцы (Dytiscidae), водяные клопы—гладыш (Notonecta), водяной скорпион (Nepa)—уничтожают икру и молодь рыб. Жук-плавунец и водяной скорпион в нерестовых прудах уничтожают молодь, а гладыш нападает и на небольших взрослых рыб. Жук-плавунец, например, может в день съесть до 8 четырехдневных рыбок. Мальков рыб поедают также крупные стрекозы.
Иглокожие (Echinodermata) используются некоторыми рыбами в пищу (например, пестрой зубаткой в Баренцевом море), а небольшая рыбка карапус (Carapus), обитающая в Средиземном море, почти в течение всей жизни живет в полости тела голотурии, питаясь на одной из стадий своего развития гонадами и водными легкими хозяина. Это сожительство не приносит вреда голотурии, поврежденные органы которой быстро восстанавливаются. Многие иглокожие, например морские звезды, морские ежи, поедая донных беспозвоночных, являются пищевыми конкурентами, а в ряде случаев и хищниками.
Земноводные (Amphibia) употребляются в пищу змееголовом, форелеокунем, сомом, щукой и др. В свою очередь лягушки уничтожают икру и молодь промысловых рыб.
Пресмыкающиеся (Reptilia) — водяные змеи, крокодилы, черепахи — полностью или частично питаются рыбой.
Некоторые птицы (Aves) поедают в водоемах головастиков, лягушек и их икру, вредных насекомых, а их экскременты являются удобрением. Нередко в прудовых хозяйствах совместно выращивают карпа и уток. В то же ,время многочисленные рыбоядные птицы (гагары, поганки, бакланы, пеликаны, цапли, чайки и др.) уничтожают большое количество взрослой рыбы и молоди.
Так, у берегов Перу бакланы ежегодно съедают около 3 млн. т анчоуса. Кроме того, являясь окончательными хозяевами некоторых опасных для рыб паразитических червей, рыбоядные птицы способствуют распространению ряда заболеваний, например, лигулеза.
Некоторые рыбы (хариус, щука, сом, гольцы, таймень и др.) употребляют в пищу мелких наземных млекопитающих (Mammalia)
— мышей и землероек. На крупных млекопитающих может нападать небольшая хищная рыба — пиранья, обитающая в реках Бразилии. Большое количество рыбы уничтожают киты и ластоногие. Например, дельфины в Черном море поедают хамсу, шпрота, мерланга, ставриду, барабулю. В пище усатых китов также часто встречается рыба (мойва, сельдь, анчоусы и др.), но основу их пищевого рациона составляют ракообразные, являющиеся кормом для рыб. Морские котики, например, в северной части Тихого океана ежегодно поедают более 2 млн. т рыбы.
Как показано, поведение рыб и их уловы зависят как от абиотических, так и биотических факторов. Поэтому изучение влияния этих факторов на рыб должно стать основой для организации рационального рыбного хозяйства и эффективности прогнозов.
Зная условия существования рыб, можно управлять биологическими процессами во внутренних водоемах, особенно в прудовых хозяйствах, повышать их рыбопродуктивность. В прудовых хозяйствах это достигается путем проведения профилактических и лечебных мероприятий против болезней и паразитов, повышения кормности водоемов за счет внесения удобрений, наиболее полного использования кормовой базы путем выращивания поликультуры. В водохранилищах и внутренних морях повышение рыбопродуктивности может быть достигнуто за счет акклиматизации рыб и водных беспозвоночных.
Дата добавления: 2020-11-18; просмотров: 2185;
Поиск по сайту
Узнать еще
- IV. Конденсация фенолов с другими органическими соединениями
- Анализ объекта оценки с позиции отражающей взаимоотношения компонентов собственности
- Аппараты для инкубации необесклеенной икры рыб (в приклеенном состоянии).
- Ассортимент и экспертиза качества рыбных консервов
- Ассортимент и экспертиза качества рыбных презервов
- Биологических особенностей половых клеток разных видов рыб
- Биотические взаимоотношения в биосфере, их закономерности. Биоразнообразие и его значение. (Модель Н.Ф. Реймерса “Волчок жизни”)
- Биотические взаимоотношения между популяциями и организмами

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
