Репродуктивные процессы в цветке
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в семенах. Цветковые являются разноспоровыми растениями. Они образуют два типа спор: микро- и мегаспоры.
Клетки бесполого размножения – микроспоры, образуются в ходе процесса, называемого микроспорогенезом в микроспорангиях, которыми являются гнезда пыльника. В результате ряда последовательных митотических делений клеток археспория, то есть образовательной ткани пыльника, возникают материнские клетки микроспор (микроспороциты), содержащие, как и все растение, диплоидный набор хромосом. Далее каждая материнская клетка в результате мейотического деления образует тетраду гаплоидных микроспор. Сформировавшаяся микроспора имеет оболочку и единственное ядро.
В ходе превращения микроспоры в пыльцевое зерно происходит два митотических деления и под защитой оболочки микроспоры возникают сначала две, далее три клетки – редуцированный мужской гаметофит. После формирования такого редуцированного гаметофита трехклеточная структура называется пыльцевым зерном (мужской особью). Пыльцевое зерно в начале развития содержит две клетки: маленькую генеративную и более крупную, часто называемую вегетативной. Микрогаметогенез крайне упрощен. Генеративная клетка делится однократно и из нее образуются две безжгутиковые мужские гаплоидные гаметы – спермии. В таком состоянии пыльцевое зерно готово к половому размножению. При попадании на рыльце пестика цветка вегетативная клетка пыльцевого зерна удлиняется и становится так называемой «пыльцевой трубкой», с мощью которой уже в семязачатке завершается оплодотворение (рис. 13.13).
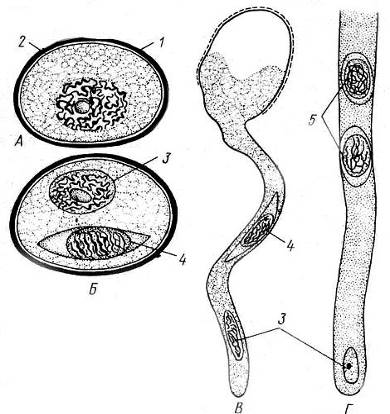
Рис. 13.13.Микроспора, пыльцевое зерно и его прорастание. А – микроспора; Б – пыльцевое зерно; В – формирование пыльцевой трубки; Г– часть пыльцевой трубки: 1 – экзина, 2 – интина, 3 – вегетативная клетка, дающая начало пыльцевой трубке, 4 – генеративная клетка, 5 – спермий.
Зрелое пыльцевое зерно окружено сложно устроенной оболочкой. Оболочка, часто называемая спородермой, состоит из двух главных слоев: внешнего, более толстого – экзины, и относительно тонкого внутреннего – интины. Экзина, содержащая особое высокомолекулярное вещество спорополленин, характеризуется необычайной стойкостью: она не растворяется в кислотах и щелочах, выдерживает температуру до 300 °С и сохраняется в фоссилизованном (окаменевшем) состоянии миллионы лет в геологических отложениях. Это позволяет вести спорово-пыльцевой анализ почв, донных отложений и осадочных толщ, определять, какие виды росли в разные геологические периоды. Экзина микроспор имеет на поверхности разнообразные скульптурные утолщения.
Особенности скульптуры важны при морфологической характеристике пыльцевых зерен отдельных таксонов. Наиболее развиты такие утолщения у энтомофильных растений. Интина менее стойка, чем экзина. Она состоит из целлюлозы и пектина и как бы облегает содержимое пыльцевого зерна.
Перенос пыльцевых зерен из тычинок на рыльца пестиков называется опылением. Для того чтобы образовался зародыш семени, должны произойти опыление и оплодотворение. Опыление впервые появляется у голосеменных, однако наибольшее разнообразие механизмов опыления и его совершенство достигаются у цветковых. Различают два типа опыления – самоопыление, или автогамию (от греческого «аутос» – сам), и перекрестное опыление, или ксеногамию (от греческого «ксенос» – чужой, «гамос» – брак). При самоопылении рыльце опыляется пыльцой того же цветка или пыльцой других цветков этой особи растения – гейтоногамия (девственное опыление). Обычно самоопыление осуществляется в распустившихся цветках, но иногда происходит в цветках закрытых, нераспустившихся (клейстогамных). В генетическом отношении все эти способы вполне равноценны.
Если перенос пыльцы осуществляется между цветками разных особей, то происходит перекрестное опыление. Оно свойственно не менее 90% видов цветковых растений. Перекрестное опыление обеспечивает обмен генами, поддерживает высокий уровень гетерозиготности популяций, определяет единство и целостность вида. Это создает широкое поле для деятельности естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, то есть делает популяции гомозиготными. Иногда это приводит к затуханию микроэволюции. Но самоопыление способствует изоляции вновь возникших в результате мутаций форм, обособляя и фиксируя их в чистых линиях. По-видимому, для эволюционного процесса оптимально сочетание самоопыления и перекрестного опыления, что чаще всего и имеет место в природе. Однако преобладает перекрестное опыление и поэтому у цветковых обычно есть специальные приспособления морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление. К ним относятся двудомность, дихогамия, гетеростилия, самонесовместимость. Наиболее надежное средство предотвращения самоопыления – двудомность, но при этом часть особей популяций (мужские особи) не дает семян. Однодомность устраняет автогамию, но не предохраняет от генетически равноценной ей гейтоногамии. Функциональная раздельнополость получила название дихогамии (от греческого «дихе» – на две части). Она проявляется в разновременном созревании пыльцы и рылец в цветках одного и того же растения. Дихогамия чаще всего встречается в форме протандрии. В этом случае раньше вызревает пыльца. При протогинии (другой форме дихогамии) происходит более раннее созревание рылец и завязей, в которых заключены семязачатки.
Иногда бывает полная физиологическая самонесовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи. Самонесовместимость встречается у покрытосеменных более широко, чем двудомность. Она зарегистрирована более чем у 10000 видов цветковых.
Явление гетеростилии состоит в том, что у некоторых видов имеются две или даже три формы цветков (находящиеся на разных особях), различающиеся по длине столбиков и тычиночных нитей. В силу этого самоопыление у этих растений в значительной мере затруднено и дает обычно ничтожное количество семян. Классическими примерами растений, характеризующихся гетеростилией, служат виды первоцветов (Primula) и болотное растение дербенник иволистный (Lythrum salicaria).
Механизмы перекрестного опыления весьма разнообразны и являются главным объектом изучения особого раздела ботаники, называемого антэкологией. Их подразделяют на два основных типа – биотическое и абиотическое опыление. Биотическое опыление осуществляется животными, абиотическое – с помощью неживых факторов внешней среды. Наибольшее значение среди механизмов биотического опыления имеет энтомофилия (от греческого «энтомон» – насекомое, «филео» – любить). Насекомые сыграли выдающуюся роль в эволюции цветка. Цветки привлекают насекомых запасом пищи: пыльцой, нектаром. Для привлечения опылителей служат яркая окраска и характерный запах цветков; некоторое значение имеет их форма. Конкретные механизмы перекрестного опыления, осуществляемого насекомыми, очень разнообразны и связаны как со строением цветка, так и с особенностями тела насекомого. Видам некоторых семейств (бобовые, губоцветные, орхидные) свойственны особые способы опыления.
Помимо насекомых существенную роль в биотическом опылении, особенно в тропиках, играют птицы (орнитофилия), летучие мыши и некоторые нелетающие млекопитающие (например, крысы). Абиотическое опыление связано с переносом пыльцы ветром (анемофилия, от греческого «анемос» – ветер), реже (у болотных и водных растений) – водой. Анемофилы – это преимущественно растения открытых пространств. У анемофильных цветковых растений, как правило, невзрачный, сильно редуцированный околоцветник и крупные, часто мохнатые рыльца с огромной воспринимающей пыльцу поверхностью. Очень часто мелкие цветки анемофилов собраны в плотные или многоцветковые соцветия, пыльца их обильна, легка, а экзина почти лишена скульптурных утолщений, обычных для пыльцы энтомофильных растений.
Второй тип спор разноспоровых цветковых растений – мегаспоры образуются в процессе мегаспорогенеза, протекающего в семязачатке. Центральная часть семязачатка – нуцеллус, представляет собой мегаспорангий цветковых растений. Семязачаток состоит из диплоидных клеток, поскольку он является частью растения-спорофита. Одна из клеток нуцеллуса является материнской клеткой мегаспор. Она редукционно делится и формирует тетраду линейно расположенных гаплоидных клеток. (рис. 13.14).
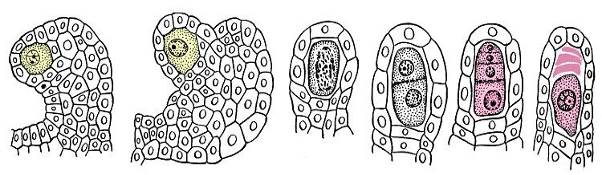
Рис. 13.14.Стадии развития мегаспор в семязачатке лобелии(Lobelia cardinalis)
Это и есть мегаспоры цветковых растений. Обычно три из четырех клеток-мегаспор в семязачатке отмирают, единственная жизнеспособная мегаспора развивается в женский гаметофит, или зародышевый мешок.
Формирование зародышевого мешка, получившее название мегагаметогенез, происходит путем трех последовательных делений ядра мегаспоры. В результате первого деления образуется двухъядерная клетка, которая сильно увеличивается в длину. Далее каждое из двух ядер делится еще дважды и возникает восьмиядерная клетка, имеющая по четыре ядра у каждого полюса. На этом деления завершаются. В центр клетки от каждого их полюсов перетекает по одному гаплоидному ядру и они сливаются в диплоидное вторичное ядро. В результате цитокинеза остальные ядра (по три у каждого полюса) формируют дочерние гаплоидные клетки. В итоге возникает семиклеточный зародышевый мешок. На микропилярном его конце обычно имеется гаплоидная яйцеклетка в окружении двух гаплоидных клеток-синергид. На халазальном конце расположены три гаплоидные клетки-антиподы. Основной объем зародышевого мешка составляет центральная диплоидная клетка (см. рис. 13.10).
Для осуществления оплодотворения необходимы два условия: зрелая жизнеспособная пыльца, попавшая на рыльце пестика, и сформировавшийся зародышевый мешок в семязачатке. Оплодотворению предшествует прорастание пыльцевого зерна. Оно начинается с разбухания зерна и выхода из апертуры пыльцевой трубки. В густой цитоплазме кончика пыльцевой трубки идут интенсивные физиологические процессы, вследствие которых несколько размягчаются ткани рыльца и столбика, в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия. В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий (нуцеллус) через микропиле семязачатка, реже – иным образом. Проникнув в зародышевый мешок, пыльцевая трубка разрывается (под действием разницы осмотического давления в ней и давления в нуцеллусе) и ее содержимое изливается в зародышевый мешок. Один из спермиев сливается с яйцеклеткой и образуется диплоидная зигота, дающая затем начало зародышу. Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра, развивающегося затем в специальную питательную ткань – эндосперм (от греческого «эндон» – внутри, «сперма» – семя). Весь этот процесс получил название двойного оплодотворения. Он впервые описан в 1898 г. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка (антиподы и синергиды) разрушаются при проникновении пыльцевой трубки. Однако при гибели яйцеклетки они могут выполнить ее функцию.
Биологический смысл двойного оплодотворения, в отличие от голосеменных, у которых гаплоидный эндосперм развивается независимо от процесса оплодотворения, состоит в том, что у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа семян этим достигается существенная экономия энергетических и пластических ресурсов.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис. Семена у апомиктичных растений образуются без оплодотворения. Апомиксис исключает генетическое расщепление, поэтому апомиктические формы образуют клоны, в пределах которых все особи имеют одинаковую генетическую и соматическую конституцию. Хорошим примером растения с апомиктическим образованием семян служит одуванчик с его поразительно высокой жизнеспособностью. Нередко у многолетних корневищных растений преобладает вегетативное размножение, а семенное ограничено.
Соцветия
Цветки могут располагаться поодиночке или группами. В тех случаях, когда они располагаются группами, образуются соцветия. Соцветием называют часть побега или систему видоизмененных побегов, несущих цветки. Соцветия обычно отграничены от вегетативной части растения. Биологический смысл возникновения соцветий – в возрастающей вероятности опыления цветков как анемофильных, так и энтомофильных растений. Несомненно, что насекомое за единицу времени посетит гораздо больше цветков, если они собраны в соцветия. Кроме того, цветки, собранные в соцветия, более заметны среди зелени листьев, нежели одиночные цветки. Многие поникающие соцветия легко раскачиваются под влиянием движения воздуха, способствуя тем самым рассеиванию пыльцы.
Соцветия свойственны большинству цветковых растений. Обычно соцветия группируются на концах вегетативных побегов, но иногда, особенно у тропических деревьев, возникают на стволах и толстых ветвях. Такое явление известно под названием каулифлории (от латинского «каулис» – стебель, «флос» – цветок). В качестве примера можно привести шоколадное дерево (Theobroma cacao). Считается, что в условиях тропического леса каулифлория делает цветки более доступными для насекомых-опылителей.
Закладываются соцветия внутри цветочных или смешанных почек. У многих растений (бузина, сирень, гиацинт) соцветие возникает как единое целое в результате деятельности одной меристемы. Любое соцветие имеет главную ось, или ось соцветия, и боковые оси, которые могут быть разветвлены в различной степени или не ветвятся. Конечные их ответвления – цветоножки несут цветки. Оси соцветия делятся на узлы и междоузлия. На узлах осей соцветия располагаются листья и прицветники (рис. 13.15).
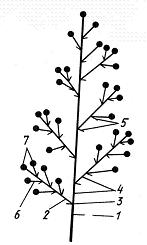
Рис. 13.15.Строение соцветия: 1 – главная ось, 2 – боковая ось (паракладий), 3 – узлы, 4 – междоузлия, 5 – прицветники, 6 – цветоножки, 7 – цветки.
Соцветие несет видоизмененные или неизмененные листья. Сильно видоизмененные листья называют прицветниками, или брактеями. Соцветие, несущее неизмененные ассимилирующие листья, является фрондозным, то есть олиственным. У брактеозного соцветия в узлах располагаются прицветники. Иногда вследствие полной редукции брактей соцветие становится эбрактеозным. У него прицветники отсутствуют. Соцветия могут быть резко отделены от вегетативной части либо (особенно в случае фрондозных соцветий) эта граница выражена неясно.
Точная характеристика соцветий необходима при анализе морфологических особенностей лекарственных растений. Поэтому классификации соцветий уделяется значительное внимание. Современная классификация соцветий, в создание которой большой вклад внес немецкий ботаник-морфолог В. Тролль (1964, 1969), довольно сложна. Поэтому для работ прикладного характера и в учебной литературе пользуются обычно различными упрощенными вариантами.
Соцветия, у которых боковые оси ветвятся, называются сложными. У простых соцветий боковые оси не разветвлены и являются цветоножками. У сложного соцветия боковые оси несут частные, или парциальные, соцветия. Считается, что сложные соцветия эволюционно возникли раньше простых. Во многих случаях простые соцветия возникали в процессе упрощения сложных, что связано с редукцией их боковых осей. Полагают, что одиночные цветки в пазухах листьев или на верхушках побегов также возникли в результат крайней редукции соцветий разного типа. Главная ось может заканчиваться верхушечным цветком – в этом случае соцветие ограничено в росте и получило название закрытого. У открытых соцветий главная ось обладает неограниченным ростом, и цветки располагаются сбоку от морфологической верхушки. У сложных соцветий верхушечными цветками могут заканчиваться главная и боковые оси, или все они имеют неограниченный рост.
У обоеполых растений соцветия несут обоеполые цветки, но у однодомных и двудомных соцветия могут быть также тычиночными, пестичными и полигамными. В последнем случае встречаются одновременно тычиночные, пестичные и обоеполые цветки. Классификация соцветий может осуществляться на основе особенностей ветвления конечных парциальных соцветий. В соответствии с этим соцветия подразделяют на два главных типа: ботриоидные (ботрические, или рацемозные) и цимоидные (цимозные). У ботриоидных соцветий (от греческого «ботрион» – кисть) характер ветвления моноподиальный. Цимоидные соцветия (от греческого «кюма» – волна по особому порядку зацветания) обязательно характеризуются симподиальным ветвлением парциальных соцветий.
Простые бoтриоидные соцветия довольно обычны, особенно у травянистых растений. Боковые оси простых соцветий не ветвятся и представляют собой цветоножки, заканчивающиеся цветками. Простые соцветия могут быть как открытыми, так и закрытыми. Возникают они, по-видимому, из сложных ботриоидных соцветий различного типа (в результате процесса редукции), а также иногда из цимоидных соцветий. Наиболее обычное простое соцветие – кисть. В кисти все цветки сидят на цветоножках, более или менее равномерно распределенных вдоль оси. Кисти встречаются у многих представителей семейства крестоцветных. Колос – производное кисти, отличающийся от нее сидячими цветками. В колос, например, собраны цветки у видов рода ятрышник (Orchis). Разновидностью колоса следует считать простую ботриоидную сережку (топольPopulus, иваSalix). Сережка обычно поникает, после цветения или созревания плодов опадает целиком вместе с осью соцветия. Чаще всего сережка несет однополые цветки (иваSalix, топольРоpulus). Почти все представители семейства аронниковых (Araceae) имеют соцветие початок, который отличается от колоса разросшейся утолщенной осью соцветия. Часто початок окружен прицветным листом – покрывалом, нередко имеющим незеленую окраску. Все перечисленные соцветия имеют нормальную развитую ось. однако существует ряд соцветий с укороченной осью. К соцветиям такого типа относятся зонтик, головка и корзинка. Зонтик – соцветие, производное от кисти, но у него все цветоножки и прицветники расположены на верхушке укороченной оси соцветия. Примерами зонтиков могут служить соцветия видов примулы (Primula) и женьшеня (Раnах). Головка представляет собой видоизмененный зонтик, у которого редуцированы цветоножки, а укороченная ось соцветия разрастается. Головка, окруженная оберткой, то есть сближенными верхушечными листьями, известна под названием корзинки. Корзинка характерна для всех представителей семейства сложноцветных.
Сложные ботриоидные соцветия могут быть как открытыми, так и закрытыми. Наиболее обычна метелка, которая представляет собой более или менее разветвленное соцветие с парциальными соцветиями ботриоидной природы. Для метелки характерно постепенное уменьшение степени разветвленности боковых осей от основания к верхушке. Классический пример метелки – метелка некоторых злаков. Метелка характерна для широко культивируемых видов сирени (Syringa) и спиреи иволистной (Spiraea salicifolia). Метелка легко видоизменяется в несколько других типов сложных соцветий, например в сложный щиток и антелу. Сложный щиток представляет собой видоизмененную метелку с укороченными междоузлиями главной оси и сильно развитыми междоузлиями боковых осей. Окончания парциальных соцветий при этом достигают уровня верхушечного (терминального) цветка. У антелы междоузлия боковых осей так сильно удлиняются, что терминальный цветок оказывается на дне воронки, образованной боковыми ответвлениями. Примером сложного щитка может служить соцветие спиреи японской (Spiraea japonica), а антелы – соцветие лабазника обыкновенного (Filipendula vulgaris). Сложная кисть представляет собой соцветие, у которого ботриоидные парциальные соцветия являются простыми кистями. Существуют разные типы сложных кистей. Наиболее обычны двойные (вероника простертаяVeronica prostrata) и тройные сложные кисти (верблюжья колючкаAlhagimaurorum). Производным от сложной кисти считается сложный колос. У сложного колоса цветки располагаются на боковых осях, то есть они сидячие, и парциальные соцветия представляют собой простые колосья. Как и сложная кисть, сложный колос может быть двойным или тройным. Сложный колос имеют большинство злаков и многие осоковые. Другое производное сложной кисти – сложный зонтик. Он известен по соцветиям представителей семейства зонтичных. Чаще всего сложный зонтик имеет боковые оси двух порядков – первого и второго. Оси первого порядка отходят от верхушки главной оси, а второго – от верхушек осей первого порядка. Парциальные соцветия сложного зонтика, таким образом, представляют простые зонтики и получили у систематиков название зонтичков. Некоторые типы ботриоидных соцветий показаны нарис.13.16.
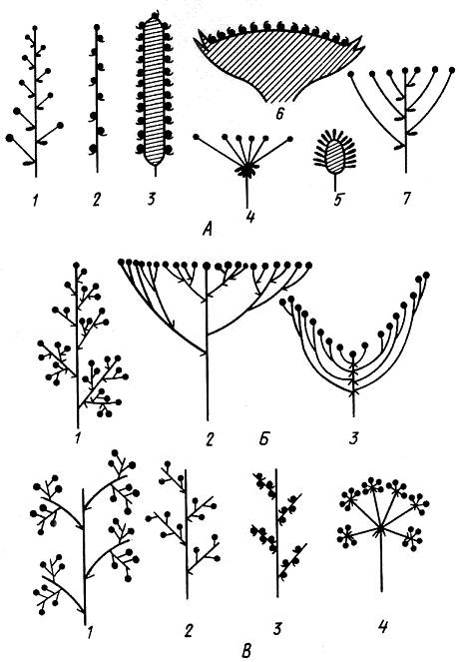
Рис. 13.16.Типы ботриоидных соцветий. А – простые ботриоидные: 1 – кисть, 2 – колос, 3 – початок, 4 – простои зонтик, 5 – головка, 6 – корзинка, 7 – щиток (4. 5, 6 – с укороченной главной осью, прочие – с удлиненной); Б – сложные ботриоидные. Метелка и ее производные:1 – метелка, 2 – сложный щиток, 3 – антела; В – сложные ботриоидные. Сложная кисть и ее производные: 1– тройная кисть, 2 – двойная кисть, 3 – двойной колос, 4 – двойной зонтик
Кроме перечисленных соцветий, существует ряд типов, у которых особенности ветвления главной оси отличаются orособенности ветвления парциальных соцветий – их называют агрегатными. Например, метелка зонтиков – метельчато ветвящееся соцветие, несущее на конечных осях простые зонтики (аралии маньчжурскаяAralia mandshurica). Метелка корзинок – метельчато разветвленное соцветие, несущее на конечных осях парциальные соцветия – корзинки. Существуют еще кисть корзинок (череда пониклаяBidens cernua), колос корзинок (сушеница леснаяGnaphalium sylvaticum) (рис. 13.17).
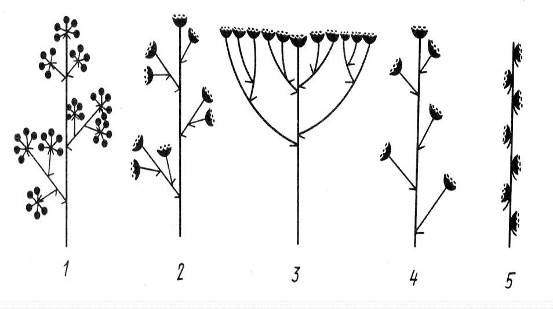
Рис. 13.17.Агрегатные соцветия: 1 – метелка зонтиков, 2 – метелка корзинок, 3 – щиток корзинок, 4 – кисть корзинок, 5 – колос корзинок.
Другую группу составляют цимоидные (цимозные) соцветия. Это обширная группа соцветий, встречающаяся так же часто, как и ботриоидные. Среди цимоидных (цимозных) соцветий выделяют два основных типа: цимоиды и тирсы (рис. 13.18).
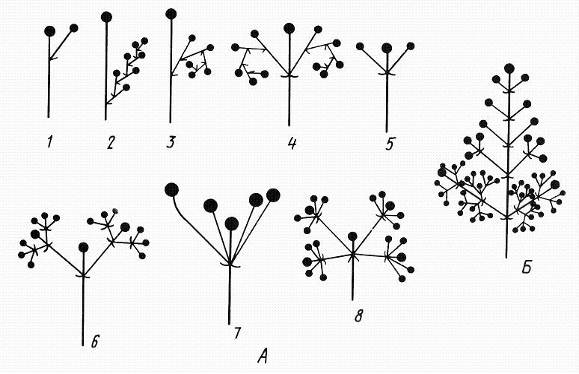
Рис. 13.18.Цимоидные соцветия. А – цимоиды: 1-3– монохазии: 1 –элементарный монохазий, 2– извилина, 3 – завиток, 4 – двойной завиток, 5-6– дихазии: 5 – дихазий, 6 – тройной дихазий, 7-8 – плейохазии: 7 – плейохазий, 8 – двойной плейохазий; Б – тирс.
Цимоиды, как правило, представляют собой упрощенные тирсы. У всех цимоидных соцветий парциальные соцветия образуются за счет симиодиального ветвления. Различают три типа цимоидов: монохазии, дихазии и плейохазии. У монохазиев под цветком, завершающим главную ось, развивается только одно парциальное соцветие или в простейших случаях единственный цветок. В соответствии с особенностями ветвления парциальных соцветий типа монохазиев принято выделять извилину, завиток и клубок. Монохазий обычен у лютиковых, в частности у лютика едкого (Ranunculus acris). Парциальные соцветия в виде завитка встречаются у большинства представителей семейства бурачниковых (Boraginaceae).
От главной оси дихазия, под завершающим ее цветком, отходят два парциальных соцветия, а в простейших случаях – два цветка. Возможны простые, двойные, тройные дихазии. Дихазий встречается у ряда гвоздичных, например видов рода звездчатка (Stellaria).
Плейохазии характеризуются строением, при котором под цветком, завершающим главную ось, развиваются три или более парциальных соцветия (или цветка). Принципиально возможны двойные, тройные и более сложные плейохазии.
Для рода молочай (Euphorbia) из семейства молочайных характерен особый тип цимоидного соцветия, получивший название циатий. Циатий состоит из верхушечного пестичного цветка и пяти тычинок, возникших вследствие крайней редукции пяти тычиночных парциальных соцветий. Циатий окружен оберткой, состоящей из листьев редуцированных парциальных соцветий.
Тирсы устроены сложнее цимоидов. Это разветвленные соцветия, причем степень разветвления уменьшается от основания к верхушке. Главная ось тирса нарастает моноподиально, но парциальными соцветиями того или иного порядка являются цимоиды. Возможны различные классификации тирсов. Тирсы, главная ось которых заканчивается цветком, называются закрытыми, в противном случае они считаются открытыми. В зависимости от степени разветвления боковых осей выделяют плейотирсы, у которых цимоиды размещаются на осях третьего и более высоких порядков; дитирсы, у которых цимоиды расположены на осях второго порядка, и монотирсы, у которых цимоиды находятся непосредственно на главной оси соцветия.
Внешнее сходство тирсов с кистью, колосом, сережкой, зонтиком или головкой позволяет говорить о кистевидном, колосовидном, сережковидном, зонтиковидном, головковидном тирсах. Тирсы легко преобразуются в другие типы соцветий в ходе редукции (уменьшения) числа боковых осей, укорочения междоузлий. В результате редукции возникают цимоиды – соцветия, по внешнему виду подобные ботриоидным, и даже одиночные цветки. Тирсы встречаются у растений очень часто. Например, тирс – соцветие конского каштана (Aesculus hippocastanum), другой пример тирса – соцветие коровяка (Verbascum) из семейства норичниковых. Тирсы различных типов представляют собой соцветия всех губоцветных. Соцветие березы – сережковидный тирс.
Многие цветки используются для получения медицинских препаратов или их компонентов. Многие цветки содержат эфирные масла. Одно из известнейших в Европе розовое масло, получаемое из лепестков махровых форм розы дамасской (Rosa damascena), розы столистной (R. сentifolia), видов жасмина (Jasminium) применяется в парфюмерии и в медицине. В медицине употребляют много цветков различных растений. Особенно популярны цветки ромашки аптечной (Matricaria chamomilla), цветки липы (Тilia cordata), кукурузы (Zeamais), цветки календулы, или ноготков (Calendula officinalis), бутоны софоры японской (Styphnolobium japonicum) – источника промышленного получения витамина Р (рутина). Ряд цветков применяют для ароматизации вин и табаков. Из них особенно известен донник лекарственный (Melilotus officinalis), чьи цветки, содержащие кумарин, добавляют во многие сорта табака. Пищевые красители получают из цветков уже упоминавшихся ноготков лекарственных, рылец культивируемого шафрана посевного (Crocus sativus) и сафлора красильного (Carthamus tinctorius). Зеленые бутоны каперсов колючих (Сарparis spinosa) маринуют в качестве острой приправы. Ценную пряность представляют бутоны гвоздичного дерева, или сизигиума ароматного (Syzygium aromaticum). Они известны пoд названием «гвоздики», что связано с формой высушенных бутонов. В тропических районах многие крупные цветки используют как овощи. Довольно широкое применение в этом качестве находят бутоны цветков некоторых видов бананов (Musa). Нектар цветков служит источником получения разных сортов меда. Наиболее ценные медоносы – разные виды липы, бобовых, астровых, фацелия пижмолистная (Phacelia tanacetifolia) и гречиха съедобная (Fagopyrum esculentum). В последние десятилетия в медицине стали использовать препараты пыльцы растений.
Дата добавления: 2016-05-30; просмотров: 4401;
Поиск по сайту
Узнать еще
- I.1.3 ТЕРМОДИНАМИЧЕСКИЙ ПРОЦЕСС. ИЗОПРОЦЕССЫ
- V. Новообразования (доброкачественные и злокачественные) и диспласти-ческие (опухолевые) процессы.
- XIX. ХОЛОДИЛЬНЫЕ ПРОЦЕССЫ
- Автогенетические процессы и половой отбор
- АДИАБАТИЧЕСКИЕ ПРОЦЕССЫ В АТМОСФЕРЕ
- Баромембранные процессы.
- Бизнес-процессы. Классификация.
- Биогеохимические циклы (биогеохимические круговороты) -циклические процессы обмена веществ между различными компонентами биосферы, обусловленные жизнедеятельностью организмов.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
