THE THIRD WEEK of LIFE. TRILAMINAR and EMBRYONIC SHIELD PERIOD
The second stage of gastrulation – immigration – occurs at the 14-th or 15th day of embryogenesis. In mammalian species, the morphological changes of the second stage of the gastrulation takes place only in the epiblast(hypoblast gives rise to the yolk sac only).
With the development of the extra-embryonic coelom, the extra-embryonic mesoderm has been divided into two zones - somatopleuric in contact with amniogenic cells, or with trophoblast, and splanchnopleuric in contact with entoderm. The early extra-embryonic membranes - amnion, chorion and yolk sac - can now be recognized (Fig.27).
By 14-15 days the second phase of gastrulation – immigration begins. In mammalian species, the morphological changes of the second stage of the gastrulation takes place only in the epiblast(hypoblast gives rise to the extra-embryonic endoderm only).
The primitive streak, the key structure of the second stage, appears in the epiblast. Epiblastic cells from the cranial end of the embryonic disk proliferate and migrate along the disc margins to its caudal end. Cellular currents converge at the disk caudal end, then turn toward the midline, and elongate back to the disk cranial end. The anterior portion of the primitive streak thickens to form the primitive knot (Hensen’s nodule). Concurrently, a narrow primitive groove develops in the primitive streak, which is continuous with a depression in the primitive knot known as the primitive pit. Since the primitive streak appears, it becomes possible to identify the craniocaudal axis of the embryo, its cranial and caudal ends, and its right and left sides.
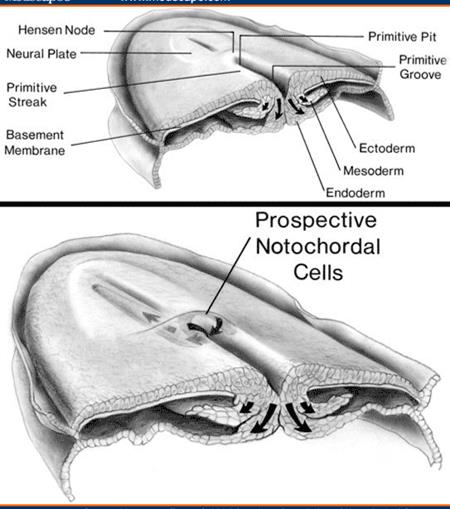
The primitive streak is a source of the embryonic mesoderm and embryonic endoderm: epiblastic cells move medially towards the primitive streak and enter the primitive groove. They lost their attachment to the rest of the epiblastic cells and migrate inwardly between epiblast and hypoblast.
The early-migrating cells are those that replace hypoblastic cells to become the endoderm. The later-migrating cells begin to spread laterally, ventrally, and cranially to form the mesoderm. As soon as the primitive streak gives rise to the mesoderm and endoderm, the cells that remain in the epiblast are referred to as the embryonic ectoderm. Thus, the gastrulation is completed: the trilaminar embryo is formed.
The subsequent processes - neural tube formation, development of mesenchyme, differentiation of intra-embryonic mesoderm on somites (dermatome, myotome and sclerotome), intermediate mesoderm (nephrogonotome) and lateral plate mesoderm (parietal and visceral layers of splanchnotome, bounding coelom cavity) proceed in the same way as they occur in birds.
The complex of axial organs includes: notochord, neural tube, mesodermal somites.
The notochord is the first to appear. The primitive pit extends into the primitive knot to form the notochordial canal. The cells migrating through this canal give rise to the notochord. The notochordial process looks like a cellular rod extending cranially from the primitive knot between the ectoderm and endoderm. The wing-like mesoderm is on each side of the notochordial process. The notochord forms a midline axis of the embryo. It is the structure around which the vertebral column forms. The notochord degenerates and disappears where it becomes surrounded by the vertebral bodies, but persists as the nucleus pulposus of the intervertebral disks. The notochord also induces the overlying ectoderm to form the neural plate, i.e., the embryonic induction of neurulation.
Neurulation is a process of the neural tube formation. As the notochord develops, the embryonic ectoderm over it (neuroectoderm) thickens to form the neural plate. On about the 18th day, the neural plate invaginates along the central axis to form the neural groove with neural folds on each side of it. By the end of the 3-rd week, the neural folds begin to move and close up with formation of the neural tube. The neural tube is then separated from the surface ectoderm that differentiates into the skin epidermis. The neural tube is the primordium of the CNS (the brain and the spinal cord). As the neural folds close, some ectodermal cells lying along and over each fold are not incorporated in the neural tube. They look like a cell mass between the neural tube and the covering ectoderm constituting the neural crest. The neural crest gives rise to the spinal ganglia and the ganglia of the autonomic nervous system, Schwann cells, the meningeal covering of the brain and the spinal cord (the pia mater and the arachnoid), pigment cells (melanocytes), adrenal gland medulla.
The mesoderm on each side of the notochord and neural tube thickens to form the longitudinal columns of the paraxial mesoderm. Each paraxial mesoderm is continuous laterally with the intermediate mesoderm, which gradually thins laterally to form the lateral mesoderm.
The paraxial mesoderm begins to divide into paired bodies called somites(32-34 in humans).This series of mesodermal tissue blocks is located on each side of the developing neural tube. Somites are subdivided into three regions: the myotome that gives rise to skeletal muscles; the dermotome that gives rise to the skin dermis; the sclerotome, from which bone and cartilaginous tissues arise.
The intermediate mesoderm differentiates into nephrogonotome that gives rise to the kidney and gonads.
Within the lateral mesoderm the space called the coelom appears dividing the lateral mesoderm into two layers: the parietal layer, the somatopleure; the visceral layer, the splanchnopleure. The coelomis then divided into the following body cavities: the pericardial, pleural, and peritoneal ones. The cells of the parietal and visceral layers give rise to the mesothelium lining these cavities. The splanchnopleure takes part in the development of the myocardium, epicardium, and the adrenal gland cortex.
Some mesodermal cells migrate and are disposed among the axial organs; they form the mesenchyme: embryonic connective tissue composing of a loose net of stellate cells embedded in gelatineous intercellular substance. The mesenchyme will give rise to the blood, all types of connective tissue, smooth muscle cells, blood vessels, microglial cells, and endocardium. Ectomesenchyme has similar properties to mesenchyme. The major difference is that ectomesenchyme is usually considered to arise from neural crest cells. Ectomesenchyme plays a critical role in the formation of the hard and soft tissues of the head and neck such as bones, muscles and most importantly the branchial arches. So, mesenchyme can have different embryonic origins (e.g., from lateral mesoderm or neural crest)
The embryonic endoderm develops into the epithelium of the gastrointestinal tract, the liver, the pancreas, the gallblader, the epithelial lining of the lungs and the urinary bladder.
Blood vessel development, i.e. angiogenesis, begins in the extraembryonic mesoderm of the yolk sac. Angiogenesis and hemopoiesis occur concurrently: the primitive blood cells differentiate from mesenchyme inside embryonic vessels. The primitive heart also arises from mesenchymal cells. It appears as paired endothelial channels called the endocardial heart tubes.
The mesenchymal cells inside the chorionic villi differentiate into blood vessels. The vessels then become connected with the embryonic vessels via the vessels in the connecting stalk. By the end of the third week, the embryonic blood begins to circulate through the capillaries of the chorionic villi. The villi containing the blood vessels are called the tertiary villi (Fig.).
Fetal circulation is entirely closed,confined to vessels within the chorionic villi. Maternalblood flow though the placenta is open. “Lakes” of maternal blood fill the intervillous space.
Early in this period, two longitudinal folds – the cranial and caudal folds – and two transverse folds appear. They convert the flat trilaminar embryonic disk into a C-shaped cylindrical embryo, and the body shape is thus established. During folding, the embryo body is separated from the yolk sac, and the primitive gut is formed. By 20-21 days the embryo detaches itself from extra-embryonic parts by the body folds formation (Fig.34-36).
Дата добавления: 2020-05-20; просмотров: 1365;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
