Организация и функции мембранных белков
Липидам в составе мембран отводят, в первую очередь, структурные свойства — они создают бислой, или матрикс, в котором размещаются активные компоненты мембраны — белки. Именно белки придают разнообразным мембранам уникальность и обеспечивают специфические свойства. Многочисленные мембранные белки выполняют следующие основные функции: обусловливают перенос веществ через мембраны (транспортные функции), осуществляют катализ, обеспечивают процессы фото- и окислительного фосфорилирования, репликацию ДНК, трансляцию и модификацию белков, рецепцию сигналов и передачу нервного импульса и др.
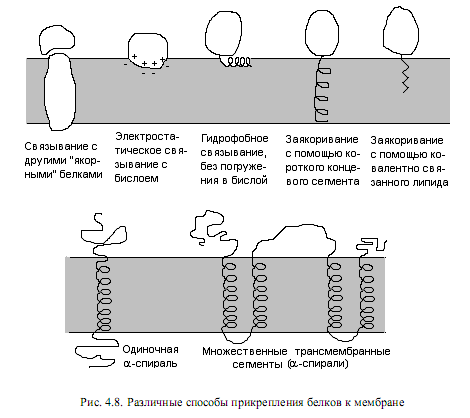
Принято делить мембранные белки на 2 группы: интегральные(внутренние) и периферические (наружные). Критерием такого разделения служит степень прочности связывания белка с мембраной и, соответственно, степень жесткости обработки, необходимой для извлечения белка из мембраны. Так, периферические белки могут высвобождаться в раствор уже при промывке мембран буферными смесями с низкой ионной силой, низкими значениями рН в присутствии хелатирующих веществ, например этилендиаминотетраацетата (ЭДТА), связывающих двухвалентные катионы. Периферические белки выделяются из мембран при таких мягких условиях, поскольку связаны с головками липидов или с другими белками мембраны при помощи слабых электростатических взаимодействий, либо с помощью гидро-фобных взаимодействий — с хвостами липидов. Наоборот, интегральные белки представляют собой амфифильные молекулы, имеют на своей поверхности большие гидрофобные участки и располагаются внутри мембраны, поэтому для их извлечения требуется разрушить бислой. Для этих целей наиболее часто используют детергенты или органические растворители. Способы прикрепления белков к мембране довольно разнообразны (рис. 4.8).
Транспортные белки. Липидный бислой является непроницаемым барьером для большинства водорастворимых молекул и ионов, и их перенос через биомембраны зависит от деятельности транспортных белков. Можно выделить два основных типа этих белков: каналы (поры) и переносчики. Каналы представляют собой туннели, пересекающие мембрану, в которых места связывания транспортируемых веществ доступны на обеих поверхностях мембраны одновременно. Каналы в процессе транспорта веществ не претерпевают каких-либо конформационных изменений, их конформация меняется лишь при открывании и закрывании. Переносчики, наоборот, в процессе переноса веществ через мембрану изменяют свою конформацию. Причем в каждый конкретный момент времени место связывания переносимого вещества в переносчике доступно только на одной поверхности мембраны.
Каналы, в свою очередь, можно разделить на две основные группы: потенциалзависимые и регулируемые химически. Примером потенциалзависимого канала является Na+ -канал, его работа регулируется изменением напряжения электрического поля. Иными словами, эти каналы открываются и закрываются в ответ на изменение трансмембранного потенциала. Химически регулируемые каналы
открываются и закрываются в ответ на связывание специфических химических агентов. Например, никотиновый ацетилхолиновый рецептор при связывании с ним нейромедиатора переходит в открытую конформацию и пропускает одновалентные катионы (подрадел 4.7 данной главы). Термины «пора» и «канал» обычно взаимозаменяемы, но под порой чаще понимают неселективные структуры, различающие вещества главным образом по размеру и пропускающие все достаточно малые молекулы. Под каналами чаще понимают ионные каналы. Скорость транспорта через открытый канал достигает 106 — 108 ионов в секунду.
Переносчики также можно разделить на 2 группы: пассивные и активные. С помощью пассивных переносчиков через мембрану осуществляется транспорт одного типа веществ. Пассивные переносчики задействованы в облегченной диффузии и лишь увеличивают поток вещества, осуществляемый по электрохимическому градиенту (например, перенос глюкозы через мембраны эритроцитов). Активные переносчики транспортируют вещества через мембрану с затратами энергии. Эти транспортные белки накапливают вещества на одной из сторон мембраны, перенося их против электрохимического градиента. Скорость транспорта с помощью переносчиков в очень сильной степени зависит от их типа и колеблется от 30 до 105 с-1. Часто для обозначения отдельных переносчиков используют термины «пермеаза», «транслоказа», которые можно считать синонимами термина «переносчик».
Ферментные функции мембранных белков. В клеточных мембранах функционирует большое количество разнообразных ферментов. Одни из них локализуются в мембране, находя там подходящую среду для превращения гидрофобных соединений, другие благодаря участию мембран располагаются в них в строгой очередности, катализируя последовательные стадии жизненно важных процессов, третьи нуждаются в содействии липидов для стабилизации своей конформации и поддержания активности. В биомембранах обнаружены ферменты — представители всех известных классов. Они могут пронизывать мембрану насквозь, присутствовать в ней в растворенной форме или, являясь периферическими белками, связываться с мембранными поверхностями в ответ на какой-либо сигнал. Можно выделить следующие характерные типы мембранных ферментов:
1) трансмембранные ферменты, катализирующие сопряженные реакции на противоположных сторонах мембраны. Эти ферменты имеют, как правило, несколько активных центров, размещающихся на противоположных сторонах мембраны. Типичными представителями таких ферментов являются компоненты дыхательной цепи или фотосинтетические редокс-центры, катализирующие окислительно-восстановительные процессы, связанные с транспортом электронов и созданием ионных градиентов на мембране;
2) трансмембранные ферменты, участвующие в транспорте веществ. Транспортные белки, сопрягающие перенос вещества с гидролизом АТР, например, обладают каталитической функцией;
3) ферменты, катализирующие превращение связанных с мембраной субстратов. Эти ферменты участвуют в метаболизме мембранных компонентов: фосфолипидов, гликолипидов, стероидов и др.
4) ферменты, участвующие в превращениях водорастворимых субстратов. С помощью мембран, чаще всего в прикрепленном к ним состоянии, ферменты могут концентрироваться в тех областях мембран, где содержание их субстратов наибольшее. Например, ферменты, гидролизующие белки и крахмал, прикрепляются к мембранам микроворсинок кишечника, что способствует увеличению скорости расщепления этих субстратов.
Белки цитоскелета. Цитоскелет представляет собой сложную сеть белковых волокон разного типа и присутствует только в эукариотических клетках. Цитоскелет обеспечивает механическую опору для плазматической мембраны, может определять форму клетки, а также местоположение органелл и их перемещение при митозе. С участием цитоскелета осуществляются также такие важные для клетки процессы, как эндо- и экзоцитоз, фагоцитоз, амебоидное движение. Таким образом, цитоскелет является динамическим каркасом клетки и определяет ее механику.
Цитоскелет формируется из волокон трех типов:
1) микрофиламенты(диаметр ~ 6 нм). Представляют собой нитевидные органеллы — полимеры глобулярного белка актина и других связанных с ним белков;
2) промежуточные филаменты (диаметр 8— 10 нм). Сформированы кератинами и родственными им белками;
3) микротрубочки (диаметр ~ 23 нм) — длинные трубчатые структуры.
Состоят из глобулярного белка тубулина, субъединицы которого формируют полый цилиндр. Длина микротрубочек может достигать нескольких микрометров в цитоплазме клеток и нескольких миллиметров в аксонах нервов.
Перечисленные структуры цитоскелета пронизывают клетку в разных направлениях и тесно связываются с мембраной, прикрепляясь к ней в некоторых точках. Эти участки мембраны играют важную роль в межклеточных контактах, с их помощью клетки могут прикрепляться к субстрату. Они же играют важную роль в трансмембранном распределении липидов и белков в мембранах.
Дата добавления: 2016-05-30; просмотров: 3281;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- II. Митохондрии (строение и функции)
- II. Организация дезинфекционных и стерилизационных мероприятий в организациях, осуществляющих медицинскую деятельность
- II. Признаки, ресурсы и функции власти.
- II. Функционально-структурная организация и программное обеспечение персонального компьютера
- III. Функции отдела по делам ГОЧС и ВМР
- III. Функции политологии. Возрастание роли политических знаний в жизни общества.
- Table 1. Функции ГИС для лесного хозяйства и лесоустройства

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
