БИОЦЕНОЗ.Видовая структура биоценоза
Популяции разных видов живых организмов, совместно заселяющие общие места обитания (биотопы), неизбежно вступают между собой и средой обитания в разнообразные отношения. Наиболее важными типами взаимосвязей являются пищевые (питание организмов одних видов другими, конкуренция за пищу и т. п.), топические (пространственное распределение, конкуренция за места поселения, убежища и т.п.) и средообразующие (формирование определенной структуры местообитания, микроклимата и пр.). Все перечисленные формы взаимоотношений осуществляются не на уровне видов (виды могут входить в состав разных биоценозов) и не на уровне отдельных особей (взаимоотношения между особями конечны, так как особь смертна).
Устойчивые, эволюционно сложившиеся взаимоотношения образуются только между популяциями видов, входящих в состав данного биоценоза. Длительное совместное существование популяций разных видов лежит в основе формирования многовидовых сообществ – биоценозов, в которых подбор видов не случаен, а определяется исторически выработанными приспособительными формами взаимоотношений. Приспособленность выражается в закономерных отношениях живых организмов друг с другом и в сходстве требований к важнейшим абиотическим факторам среды.
Биоценоз. Представляет собой эволюционно сложившуюся форму организации живых организмов, многовидовую биологическую систему на том или ином пространстве. В ее состав входят популяции организмов разных таксонов, отличающихся по своим экологическим, морфологическим и физиологическим свойствам и связанных многими видами биотических отношений как между собой, так и с окружающей абиотической средой. Именно эти связи удерживают организмы биотического сообщества друг с другом, определяют его целостность и способность к длительному самоподдержанию — гомеостазу.
Термин «биоценоз» (от греч. bios - жизнь, koinos - общий) ввел в экологическую литературу в 1877 г. немецкий зоолог К. Мебиус (1825–1908) в книге «Устрицы и устричное хозяйство», в которой специальную главу посвятил обсуждению проблемы устричной банки как биологического сообщества, или биоценоза.
Биоценоз- исторически сложившаяся группировка популяций разных видов растений (фитоценоз), животных (зооценоз), грибов (микоценоз), микроорганизмов (микробоценоз), заселяющих относительно однородное жизненное пространство и возникшее на основе осуществления биогенного круговорота веществ (рис. 1).
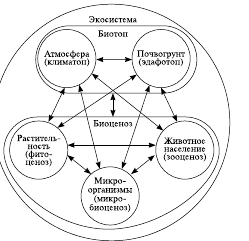
Примерами биоценозов являются еловый, сосновый, смешанный лес, луг и т. п.
Каждый биоценоз развивается в пределах биотопа – однородного пространства, которое характеризуется определенным сочетанием абиотических факторов, таких как количество приходящей солнечной энергии, температура, влажность, подвижность воздушной или водной среды, химический и механический состав почвы, ее кислотность, рельеф местности и др. Биотоп является общим местом обитания всех живых организмов биоценоза, необходимым условием его существования. От свойств биотопа зависит, какие группировки видовых популяций биоценоза могут его заселить, взаимодействуя с ним. Изменения свойств биотопа приводят к изменению структуры зависящего от него биоценоза вплоть до его деградации и разрушения.
Видовая структура биоценоза. Биоразнообразие. Каждый биоценоз можно описать, основываясь на совокупности составляющих его видов. Видовой состав различных биоценозов различается в большей или меньшей степени, что обусловлено различным географическим положением экосистем, а также средой жизни (водной, наземно-воздушной, почвенной).
Попытки оценить степень биоразнообразия биоценозов впервые были предприняты биогеографами XVIII-XIX вв. с целью разделения поверхности планеты по степени своеобразия флоры и фауны.
Термин «биоразнообразие» впервые применил Г. Бейтс в работе «Натуралист на Амазонке» (1892), в которой дал описание встреченных им 700 видов бабочек во время часовой прогулки.
В последние десятилетия термин «биоразнообразие» стал одним из самых распространенных в научной литературе, природоохранном движении и международных связях. В ходе научных исследований показано, что необходимым условием нормального стабильного функционирования экосистем и биосферы в целом является достаточный уровень природного разнообразия. В настоящее время биоразнообразие рассматривается как основной параметр, характеризующий состояние надорганизменных систем.
В ряде стран именно биоразнообразие региона выступает в качестве основы экологической политики государства, стремящегося сохранить свои биоресурсы, чтобы обеспечить устойчивое экономическое развитие.
Понятие «биологическое разнообразие» активно используется более полувека. За это время много сделано для понимания самого явления и разработки методов его измерения.
В 1972 г. на Стокгольмской конференции ООН по окружающей среде экологи впервые сумели убедить политических лидеров стран мирового сообщества в том, что охрана живой природы должна стать приоритетной при любой деятельности человека на Земле. В 1992 г. в Рио-де-Жанейро на конференции ООН была принята Конвенция о биологическом разнообразии. Ее подписали, а позднее и ратифицировали в 180 странах мира.
Международная программа исследования биоразнообразия включает три основных уровня: генетический, таксономический и экологический.
Генетическое разнообразие – поддержание генетической гетерозиготности, полиморфизма, которые вызваны адаптационной необходимостью в природных популяциях и представлены наследуемым разнообразием внутри организмов и между популяциями. Новые генетические вариации возникают в результате генных, хромосомных, геномных мутаций, а также через рекомбинацию генов. Каждая особь, а тем более вид, – уникальны. Вымирание лишь одного дикого вида означает безвозвратную потерю от тысячи до сотен тысяч генов. Основной резервуар генетических ресурсов – природные экосистемы, которые в настоящее время сильно изменены или разрушены человеческой деятельностью. Уменьшение генотипического разнообразия ставит на грань риска возникновение возможных будущих адаптаций в экосистемах.
Экологическое (экосистемное) разнообразие понимают как разнообразие биоты, обитающей на разнообразии биотопов, за счет взаимодействия которых формируются отличительные характерные особенности экосистем как малых, так и огромных размеров (биомов). Чаще всего оно оценивается через разнообразие видового компонента (биоты).
Таксономическое (видовое) разнообразие нередко рассматривают в качестве синонима богатства видов, которое понимают как число видов в конкретном местообитании, биотопе или сообществе, биоценозе.
В 1960 г. Р. Уиттекер предложил уточненную шкалу оценки видового разнообразия:
a-разнообразие – видовое разнообразие внутри местообитания или одного сообщества (биоценоза);
b-разнообразие – разнообразие между местообитаниями или разными сообществами (биоценозами);
g-разнообразие – разнообразие в обширных регионах биома, острова, континента.
В 1979 г. Д. Крюгер и Ф. Тейлор дополнили шкалу, введя
d-разнообразие, которое определяется изменениями климатических факторов, что выражается в смене растительных зон и т. д.
Измерение и оценка биоразнообразия. Контроль над биоразнообразием и его сравнение требуют его количественного измерения. Оценка величины биоразнообразия имеет важное прикладное значение, так как:
1) позволяет контролировать сохранение генетического потенциала;
2) дает представление о состоянии экосистем на определенной территории;
3) служит основой для разработки системы менеджмента популяций отдельных видов.
Параметры и методы оценки a-разнообразия. Компонентами биоразнообразия внутри сообщества являются два показателя: видовое богатство и относительное обилие видов.
Видовое богатство – число видов в отдельном сообществе или биоценозе. По этому показателю биоценозы (или отдельные сообщества) бывают весьма богатыми, насыщенными большим числом разнообразных видов. Таковы, например, тропические влажные леса (джунгли), на 1 га которых число видов только деревьев может насчитывать до 200, в то время как биоценоз соснового леса в условиях Беларуси может включать максимум до 10 видов древесных пород. Огромным видовым богатством обладают нетронутые антропогенным воздействием биоценозы коралловых рифов. И напротив, видовое богатство биоценозов высоких широт (например, тундры) значительно беднее, так как лишь немногие виды приспособились к жизни в условиях длительного периода экстремально низких температур зимнего периода года и короткого вегетационного сезона.
Однако знание числа видов в сообществе не дает полной информации о их роли в нем, так как ни одно сообщество не состоит из видов равной численности. Обычно немногие виды преобладают, т. е. доминируют, большинство же видов – малочисленны. Отсюда вытекает важность оценки относительного обилия видов, или выравненности, – степени равномерности распределения видов по их обилию в сообществе. Выравненность – важнейший показатель структуры сообщества. Значимость его оценки продемонстрируем на примере двух гипотетических упрощенных сообществ, которые имеют по 10 видов с их суммарной численностью в каждом, равной 1000 экземплярам.
В первом сообществе популяция каждого вида имеет равную численность по 100 экземпляров; во втором – один из видов имеет численность 910 экземпляров, а остальные девять видов по 10 экземпляров каждый. Таким образом, в первом сообществе выравненность максимальна, так как каждый вид имеет равное обилие; во втором – минимальна, поскольку один вид доминирует. Следовательно, во втором сообществе функциональная роль одного из видов будет определяющей, главенствующей. Для большинства сообществ свойственно наличие доминирующих (преобладающих), второстепенных, редких и случайных видов. Видовое богатство биоценоза или сообщества определяется главным образом совокупностью редких видов, так как их большинство.
Среди доминантных видов, преобладающих по численности, биомассе или продукции, особую роль играют эдификаторы, которые в ходе своей жизнедеятельности в наибольшей степени создают среду для других членов сообщества. Их называют созидателями, строителями биоценоза. Примером растений-эдификаторов в биоценозах являются: в лесах – сосна, ель, дуб, в степях – ковыль, на низинных болотах – осоки, на верховых – сфагновый мох. Своей жизнедеятельностью они так изменяют условия среды, что она становится пригодной лишь для обитания организмов определенных видов, приспособленных к жизни в измененных условиях. Изъятие доминанта из биоценоза сказывается не только на жизни всей биоты, но и на физической среде.
Для получения более полной картины о видовой структуре в сложном сообществе строят график последовательности каждого вида (ранг вида) от его обилия, используя данные по их численности или биомассе. На рис. 2 представлены примеры кривых «ранг – обилие» в трех сообществах.
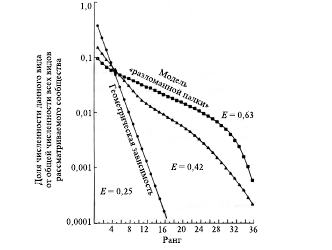
Рис. 2. Примеры рангового распределения обилий для трех гипотетических сообществ. Указаны
соответствующие каждой кривой значения равномерности распределения (E, по Симпсону)
Кривая 1-го типа (Е = 25) свидетельствует о высокой степени доминирования и низкой выравненности, а также о низком видовом богатстве. В таком сообществе имеются свободные экологические ниши и возможно вселение новых видов. Кривая 2-го типа (Е = 42) указывает меньшую степень доминирования, большую выравненность и большее видовое богатство по сравнению с 1-м типом. В таком сообществе все экологические ниши заняты, но не перекрываются. Их неперекрывание обеспечивается межвидовой конкуренцией и территориальным поведением. Кривая 3-го типа (Е = 63) иллюстрирует высокую степень выравненности и большое видовое богатство. В таком сообществе все экологические ниши заняты и перекрываются между собой, поэтому вселение новых видов сюда невозможно. Таким образом, чем круче идет кривая (1 > 2 > 3), тем больше доминирование и меньше выравненность (1 < 2 < 3).
На основании результатов оценки видового богатства и выравненности в сообществах разных природных зон планеты было сформулировано правило биогеографического градиента биоразнообразия (правило Уоллеса): видовое богатство и выравненность в биоценозах возрастают в направлении от высоких к низким широтам.
Выделяют две группы индексов биоразнообразия.
1. Простые индексы видового разнообразия (d), в основе которых лежат различные сочетания числа выявленных видов (S) и общей численности особей всех видов (N) сообщества. Их существует несколько. Приведем расчеты биоразнообразия с использованием двух индексов на примере сообщества птиц, насчитывающего 17 видов и представленных 149 особями:
индекс Маргалефа (1958)
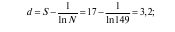
индекс Менхиника
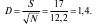
Достоинством индексов этой группы является легкость расчета.
2. Индексы неоднородности — несколько более сложные для расчета индексы, учитывающие одновременно оба параметра биоразнообразия: видовое богатство и выравненность:
индекс разнообразия Симпсона (C)
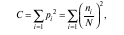
де ni — значимость отдельных видов; pi — относительные значимости для этих же видов; N — сумма значимостей для всех видов биоценоза;
индекс Шеннона — Винера (Н)
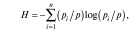
где p — общая численность (биомасса) каждого i-го вида.
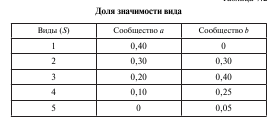
Методы оценки b-разнообразия. К наиболее простым и широко применяемым способам измерения степени сходства видового состава между разными сообществами принадлежат коэффициенты общности(КО) Жаккара и процентного сходства (ПС) Съеренсена.В качестве примера возьмем два простых сообщества, насчитывающих пять видов, из которых три являются общими. Для каждого вида приведена его доля значимости в сообществе (табл. 1).
КО Жаккара 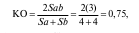
где Sa — число видов в сообществе а; Sb — число видов в сообществе b; Sab — число видов общих в обоих сообществах.
Таким образом, КО Жаккара выражает только степень видового сходства на основании присутствия или отсутствия видов в обоих сообществах. В данном примере КО составляет 75 %.
Расчет ПС основан на сумме различий значимости (доли) видов, т. е. отражает степень сходства количественного участия видов в сообществах. С учетом выравненности ПС для данных сообществ составляет 60 %, что на 10 % меньше, чем рассчитанное по КО Жаккара.
Видовое разнообразие и устойчивость биоценозов. Поскольку биоценозы представляют собой совокупности взаимодействующих популяций разных видов, то их устойчивость должна определяться относительной стабильностью самих популяций. Экологи, изучающие динамику численности популяций и факторы, ее определяющие, склонны полагать, что не существует единого механизма управления ее динамикой. Стохастизм и регуляционизм, не являются альтернативными механизмами, влияющими на динамику популяций. Как сама плотность популяции, так и изменение среды обитания могут воздействовать на величины рождаемости и смертности и приводить к повышению или понижению численности. Популяции многих видов, сталкивающиеся с периодами неблагоприятных условий среды, обладают буферными свойствами, защищающими популяцию от возрастающих потерь особей. К буферным свойствам видовых популяций относятся сохранение генетически лучше приспособленных особей в неблагополучные периоды внешней среды и обеспечение выживания популяции в последующем за счет их размножения. К буферным механизмам относятся также образование хорошо защищенных покоящихся стадий в виде спор, цист, геммул и др., которые менее чувствительны к неблагоприятным условиям среды. За счет их развития в благоприятных условиях среды популяция как форма существования вида сохраняется.
Существует мнение о том, что стабильность биоценоза – результат наличия большого видового богатства. И действительно, многовидовой биоценоз тропического леса (джунглей) весьма стабилен, в то время как маловидовые биоценозы Арктики относительно нестабильны, в них отмечается цикличность флуктуаций популяций многих составляющих их видов. Известный эколог Р. Уиттекер (1980), анализируя огромный массив накопленных данных, пришел к сомнению в том, что сложность биоценоза ведет к стабильности. Многие фактические данные свидетельствуют о том, что относительная стабильность биоценозов является результатом функции популяций отдельных видов и небольших групп взаимодействующих видов, а не видового богатства всего биоценоза. Неизменность среды обитания может обеспечить выживание многих видов, влияющих друг на друга в условиях сложного биоценоза. Некоторые виды в таком случае имеют относительно стабильные популяции (вследствие зависимости от плотности), однако сложный биоценоз может оказаться неустойчивым, если его среда нарушается. Результаты исследований показывают, что не сложность биоценоза приводит к его стабильности, а стабильность среды позволяет развиваться сложным биоценозам. Относительная стабильность природных биоценозов есть результат включения в них видов, которые приобрели в ходе эволюции механизмы, ограничивающие их флуктуации (свойства буферности и зависимости от плотности).
Биоценозы представляют собой сочетания видовых популяций, которые по разному реагируют на флуктуации условий среды и по-разному взаимодействуют друг с другом. Каждый вид имеет ограниченное число связей с другими, что приводит к отчетливой организации биоценоза. Эта организация проявляется в уникальной способности абсорбировать, или «поглощать», воздействия разного рода, что способствует выживанию биоценоза, поддержанию его жизнеспособности. Хорошим примером важности знания того, кто с кем связан и как это скажется на жизнедеятельности биоценоза, служат результаты проведенных исследований по изучению ихтиофауны Северного моря.
Исследования показали, что крупные морские рыбы Северного моря (сельдь и скумбрия) были почти полностью истреблены в результате перелова человеком. В то же время наблюдался рост численности глубоководных рыб. На первый взгляд пространственное разделение этих двух групп (одна живет на глубине, а другая у поверхности) делает данное явление неожиданным. В действительности же, как показали исследования, исчезновение сельди и скумбрии ослабило межвидовую борьбу за существование между малыми морскими рыбами (песчаным угрем, норвежским сомиком) и мальками придонных рыб. Поскольку эти виды, в противоположность сельди, мигрируют между верхними и нижними слоями, то они образуют своеобразный транспортер, приносящий энергию и пищу донным и придонным рыбам. С уничтожением главного конкурента и врага (сельди) этот транспортер смог переносить на глубину больше ресурсов, в результате чего и возросли популяции придонных обитателей. Таким образом, имеется некоторое число определенных типов связей, которые могут вызывать разнообразие последствий.
Неправомерно считать все связи одинаково существенными. Можно было бы ожидать, что исчезновение больших морских рыб должно повлиять на многих других пелагических рыб, а также их экологических соседей – морских беспозвоночных, являющихся их пищей. Однако оказалось, что высвобожденная энергия устремилась через один конкретный канал к относительно удаленной части пищевой сети.
Длительное существование видов было бы совершенно невероятным, если бы их судьба напрямую зависела от всех остальных видов в системе. Как выяснилось, экосистемы обнаруживают такую структуру связей, которая приводит к существованию подсистем, сильно связанных внутри себя, но незначительно взаимодействующих между собой. Поэтому биоценозы демонстрируют высокую способность к выживанию за счет того, что нарушение внутри одной подсистемы не обязательно приводит к разрушению всей системы. Вследствие наличия слабых связей между подсистемами оставшиеся связи помогают всей системе пережить период, достаточный для самовосстановления утраченных. Характер связей с другими подсистемами может оставаться неизменным, в то время как внутри подсистемы могут происходить кардинальные изменения. Одни виды постепенно заменяются другими, выполняющими те же самые функции, что позволяет биоценозу сохраниться.
2. ЭКОЛОГИЧЕСКАЯ НИША
История концепции ниши. Положение конкретного вида в многовидовом биоценозе определяется, с одной стороны, набором требований к абиотическим условиям, а с другой – комплексом связей с популяциями других видов и формой участия в общих функциях биоценоза. Длительное существование популяций разных видов в составе биоценоза привело к эволюционному становлению такой системы взаимоотношений между ними, при которой каждый из них пространственно и функционально занимает в нем определенное положение. Положение вида в биоценозе рассматривается как его экологическая ниша.
Впервые понятие «ниша» ввел американский зоолог Дж. Гриннелл (1917) для обозначения самой мелкой единицы распространения вида. Он делал акцент на местообитании, пространстве, где зафиксирован вид. При этом он подразумевал, что ниши разных видов не перекрываются, и таким образом определял потенциальный характер распространения отдельного вида в отсутствие взаимодействий с другими видами.
Он сформировал представление о пространственной нише вида. Такая трактовка ниши вида соответствовала аутэкологическому подходу в экологических исследованиях того времени, мало связанному с изучением экосистемного уровня организации жизни.
Ч. Элтон в труде «Экология животных» (1927) включил в понятие ниши пищевые потребности организмов и добавил, таким образом, к пониманию этого понятия функциональный аспект – трофическую нишу, описывая ее как место организмов популяции определенного вида в биотической среде в смысле их пищевых связей и взаимодействий с конкурентами за пищу. Изучение роли пищевых связей между организмами разных видов в биоценозе привело Ч. Элтона к формулированию концепции трофических уровней экологической пирамиды.
Одновременно с разработкой концепции ниши развивалась связанная с ней концепция конкурентного исключения. Проведенные в период между
20-40 гг. ХХ в. математические расчеты А. Лотки и В. Вольтерры, а также лабораторные эксперименты Г. Гаузе (на инфузориях) и Т. Парка (на мучных хрущаках) показали, что при взаимодействии между особями популяций двух видов, потребляющих сходный пищевой ресурс, имеет место конкурентное исключение. Следствие из этого: если два вида сосуществуют, то между ними должно быть какое-то экологическое различие, что позволяет им занимать свою, особую нишу.
Анализ огромного накопленного к тому времени материала позволил американскому экологу Дж. Хатчинсону (1958) обосновать концепцию многомерности экологической ниши. Он считал, что нишу следует определять с учетом всего диапазона абиотических и биотических переменных среды, к которым должен быть адаптирован данный вид и под действием которых популяция способна жить и возобновляться бесконечно долгое время. В идеале каждую такую переменную можно рассматривать как некий градиент, на котором у каждого вида имеется свой диапазон активности и устойчивости. Примером может служить освещенность в лесу, которая убывает от вершин к нижнему пологу по мере того, как растения преграждают путь свету. Эволюция видов идет в соответствии с этим градиентом, и каждый из них адаптируется к разным диапазонам освещенности.
В некоторых бореальных лесах это приводит к идеальной структуре, основанной на распределении растений по высоте и объединяющей 5-7 вертикальных ярусов, в число которых входят: высокие деревья лесного полога, более низкие деревья, высокие и низкие кустарники и травы, мох, практически не возвышающийся над уровнем земли.
Градиент каждого фактора среды можно представить как некоторое измерение пространства. Если данная ниша имеет n значимых измерений, то ее можно описать в терминах n-мерного пространства, или гиперпространства. Мерность ниши – число факторов в нише. Гиперпространство ниши – пространство, которое нельзя себе представить физически в силу того, что число осей (признаков ниши) превышает три. Таким образом, Дж. Хатчинсон под многомерной экологической нишей понимал nмерный гиперобъем, охватывающий полный диапазон условий, при которых организмы популяции определенного вида могут успешно воспроизводить себя и длительно поддерживать свое существование.
Трактовка ниши вида по Дж. Хатчинсону включает не только его местообитание, но и то, что он делает, т. е. как он преобразует энергию, каково его поведение, как он реагирует на многочисленные параметры изменчивости абиотической и биотической сред и изменяет их. Ю. Одум (1975) приводит такую аналогию: местообитание – адрес организма, а экологическая ниша – его профессия. Поэтому знание местообитания – только начало исследования вида. Чтобы определить его положение в природном сообществе, эколог должен располагать сведениями о его питании и источниках энергии, скоростях метаболизма и роста, его влиянии на другие организмы, с которыми он вступает в контакт, его пределах, в которых он влияет и способен влиять на важные процессы в экосистеме.
Дж. Хатчинсон различает два состояния видовой ниши: фундаментальную нишу, охватывающую все множество оптимальных условий, в которых вид может обитать в отсутствие конкуренции и врагов, и реализованную нишу, или тот фактический комплекс реальных условий, в которых этот вид обычно существует в природных экосистемах. Поэтому сам по себе процесс дифференциации ниш и их «упаковки» – результат не только генетически обусловленных свойств живых организмов и их фенотипической реализации, но и взаимоотношений с прочими компонентами экосистемы. Реализованная ниша, естественно, всегда меньше фундаментальной.
Ниша как спектр использования ресурсов. Ширина (размер) ниши имеет важное значение для теории ниши, поскольку понимание факторов, которые ее контролируют, приведет к более глубокому пониманию причин видового разнообразия. При многомерном подходе ширина ниши определяется как общая сумма всего разнообразия ресурсов, используемых популяцией вида. Из-за невозможности оценки всего разнообразия используемых ресурсов ее описывают лишь по одной или нескольким важнейшим осям ниши, характеризующим наибольшую активность вида.
Каждый из видов может приспособиться или быть устойчивым к некоторому диапазону каждой переменной ниши. Верхние и нижние пределы этих переменных для вида отмежевывают ту часть пространства ниши, которую он в состоянии занимать.
Это хорошо и наглядно демонстрируется на ареалах видов, показывающих границы их распространения, где главным образом температура и влажность являются определяющими факторами.
Большинство видовых популяций не обитает в своей потенциальной фундаментальной нише, а вследствие взаимодействий с другими организмами (хищничество, конкуренция) занимает меньшую по размерам реализованную нишу. Ниши видов, входящих в биоценоз, не представляют собой дискретные единицы, они в большей или меньшей степени перекрываются между собой. Расширение или сужение ширины ниши может приводить к повышению или снижению численности взаимодействующих популяций вплоть до исключения вида с низкой активностью, что и наблюдается в природе.
В результате длительной эволюции популяции виды, входящие в состав биоценоза и делящие градиент ресурса, постепенно уменьшали ширину своей ниши, что приводило к экологической дивергенции ниш видов (рис. 3) и возрастанию биоразнообразия.
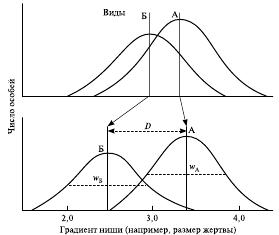
Рис. 3. Дивергенция экологических ниш у двух видов (схема): вверху — два вида с широко перекрывающими друг друга параметрами ниш, внизу — виды, которые существенно различаются по среднему параметру используемых ниш
Разнообразие ниш среди видов биоценоза возрастает благодаря невыгодности для одного или обоих видов отбора в направлении прямой конкуренции по сравнению с преимуществом отбора в сторону дифференциации ниш, дающей более надежное обеспечение ресурсами разного типа.
Например, дифференциация ниш у трех видов ящериц происходит по градиенту ресурса – размеру жертвы. Если два вида А и Б могут делить между собой градиент ресурса, то это же могут сделать и несколько видов одновременно. Группу видов, тесно связанную друг с другом своими нишами в пределах биоценоза, называют гильдией. Виды, формирующие последовательности ниш, могут эволюционировать в направлении определенных количественных связей друг с другом. Величина перекрывания популяций для конкурирующих видов ограничена. Если на градиент ресурса добавляется новый вид, ширина ниши уже имевшихся здесь видов будет сокращена.
Наблюдения за последовательностью ниш позволяют сделать некоторые заключения:
1) виды эволюционируют в направлении специализации к разным частям градиента ресурса, что уменьшает конкуренцию между ними;
2) в любой период виды стремятся к равномерному распределению адаптивных центров по градиенту ресурса;
3) последовательности ниш могут удлиняться за счет внедрения новых видов между старыми (коренными), что сокращает ширину ниш ранее существовавших видов;
4) имеются, вероятно, пределы количества видов, которые могут «упаковаться» в последовательность видов данного градиента ресурса.
Вселение в биоценоз новых видов. Это может происходить естественным путем.
Например, многих мелких насекомых и клещей переносят на огромные расстояния перелетные птицы, иногда сильные ветра. Одиночный вид-вселенец, если он находит в новой среде подходящий для него комплекс необходимых условий, постепенно встраивается в систему взаимосвязей новой экосистемы, иногда в силу большей конкурентоспособности может вытеснить один из аборигенных видов, а иногда находит свою уникальную экологическую нишу.
С развитием цивилизации все чаще причиной вселения новых видов становится деятельность человека. Человек расселил практически по всем континентам культурные растения и домашних животных. Вместе с ними в новые места попало большинство их вредителей и паразитов. Так, например, картофель ввезен в Европу из Америки еще в XVII в., а его вредитель – колорадский жук – «догнал» его в середине XX в. В одних случаях, как это было в Америке с мустангом, потомком одичавших лошадей, привезенных испанскими конкистадорами, вид-вселенец удачно вписался в структуру местных экосистем. В других, как в случае завоза в Австралию кроликов, он стал серьезным конкурентом многих видов аборигенной фауны.
В настоящее время интенсивно расселяется по пресным водоемам Европы, главным образом с участием человека, пресноводный двустворчатый моллюск дрейссена. Массовое размножение этого моллюска-фильтратора, поедающего мелких планктонных организмов, приводит к значительному снижению численности многих аборигенных видов моллюсков, рыб и их молоди, также питающихся этим же кормом. Весь биоценоз под влиянием фильтрационной деятельности дрейссены изменяется за счет подавления видов, связанных с планктоном, и постепенно превращается преимущественно в биоценоз бентосного типа за счет перекачки органического вещества в придонные области.
Аналогичная ситуация происходит в экосистеме Черного моря под влиянием гребневика мнемиопсиса, личинки которого в конце 1970-х гг. попали туда с балластными водами судов, приходивших из Америки. Сильно размножившись и поедая планктон, популяция вселившегося гребневика сильно сократила пищевую базу аборигенных промысловых рыб, уловы которых сократились в несколько раз.
Ниша — свойство вида или биоценоза? Ниша представляет собой свойство данного биоценоза и вне его лишена смысла. Ниши создаются абиотическими и биотическими компонентами экосистемы. Они заполняются в результате адаптации видов на протяжении некоторого периода эволюционного времени. Поэтому следует ожидать, что в экосистемах со сходными условиями среды биоценозы должны быть построены сходным образом и содержать одну или несколько в основном сходных ниш. Адаптации популяций, занимающих такие ниши в этих независимо сложившихся сообществах, также должны быть сходными, даже если сами виды совершенно неродственны между собой. Это явление называется экологической эквивалентностью или конвергентной эволюцией, а виды, занимающие сходные ниши – викарирующими. Наличие конвергентной эволюции и викарирующих видов служит доводом в пользу того, что ниша порождается экосистемой. Примерами могут служить присутствие суккулентов в пустынях США, представленных кактусами, экологически почти идентичными растениям из семейства молочайных, распространенных в пустынях Южной Африки. Виды злаковых трав, которыми питаются травоядные, внешне очень сходны по всей планете, хотя конкретные виды, роды и даже семейства могут быть приурочены строго к определенному материку или биогеографической области в его пределах. Кенгуру в Австралии – экологический эквивалент бизона и вилорогой антилопы в Северной Америке.
Жизнь в определенных условиях среды создает в процессе эволюции комплекс видовых приспособлений, обеспечивающих успешное выживание и воспроизведение видов в данной экологической нише. В этом случае говорят о жизненных формах различных видов. Жизненные формы организмов, занимающих одинаковые экологические ниши, могут быть причиной морфологического сходства представителей таксономически неродственных видов. Классическими примерами такого сходства являются «ихтиозавр – тунец – дельфин» (быстро плавающие обитатели водных пространств); «крот –медведка» (обитатели почвы).
Датский ботаник К. Раункиер (1934) организовал различные виды растений в пять групп жизненных форм, взяв за основу их выделения положение почек или верхушечных побегов во время неблагоприятных сезонов года: у фанерофитов почки возобновления расположены высоко (более 30 см) над землей; у хамефитов – невысоко или у самой поверхности земли; у гемикриптофитов – на почве или в ее поверхностном слое, у криптофитов – в почве или под водой; у терофитов возобновление идет только семенами. Основанная на этих признаках система жизненных форм неплохо отражает суммарную приспособленность к перенесению неблагоприятных условий наиболее уязвимой части в выживании организмов.
Сложность общей структуры биоценоза (количество ниш в нем) существенно зависит от степени гетерогенности абиотической среды (биотопа). Чем разнообразнее условия, тем большее количество отличающихся по биологическим особенностям видов могут освоить данный биотоп, тем большее видовое богатство будет в нем. Как уже было сказано, повышение биоразнообразия биоценоза связано с уменьшением ширины ниши и ростом экологической специализации видов. Немецкий эколог А. Тинеманн в 30-е гг. ХХ в. обобщил эту закономерность, отметив, что в благоприятных условиях среды высоко число видов, но каждый из них представлен относительно небольшим числом особей; в неблагоприятных условиях это соотношение меняется на обратное («правило числа видов и числа особей»). Известно выражение А. Уоллеса о том, что в тропических лесах легче за день поймать по одной бабочке ста видов, чем сто бабочек одного вида.
Следует отметить, что «нишевая» структура биоценозов открывает возможность некоторой свободы в формировании сообщества: биологически сходные виды могут в известной степени замещать друг друга в разных экосистемах. Степень «жесткости» видового состава биоценозов, возможность замещения сходных ниш разными видами зависит от возраста данной экосис<
| <== предыдущая лекция | | | следующая лекция ==> |
| Цикловая (кадровая) синхронизация. | | | Подумайте какие разделы могут быть в юридической психологии. |
Дата добавления: 2022-04-12; просмотров: 152;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
