Обмін білків в організмі.
Протягом всього життя в організмі відбуваються одночасні руйнування і біосинтез клітин і тканин. Ці протилежні, але тісно зв'язані між собою процеси – асиміляція і дисиміляція – складають основу життя. Отже, в організм повинні постійно надходити речовини, необхідні для побудови нових кліток. Головна роль у цьому належить білкам, тому що ні вуглеводи, ні жири не можуть їх замінити в утворенні основних структурних елементів органів і тканин. Серед різноманітних перетворень, властивих живій матерії, основне місце займає білковий обмін.
У зв'язку з тим, що білки є азотвмісними речовинами, одним з методів, що характеризують стан білкового обміну в організмі, може бути визначення балансу нітрогену. У здорової людини при нормальному харчуванні відзначається стан білкової рівноваги, коли надходження нітрогену компенсує його витрати. При негативному азотистому балансі кількість виведеного нітрогену перевищує його кількість, що надходить. Такий стан може спостерігатися при порушенні діяльності травної системи, білковому голодуванні і т п.
Позитивний азотистий баланс буває в тих випадках, коли кількість виведеного азоту менше того, що надходить у складі білків. Це характерно для зростаючого організму, при вагітності, при підвищенні активності процесів біосинтезу білка (наприклад, при фізичних навантаженнях).
Для синтезу білків в організмі необхідні різні амінокислоти. Деякі з них, що утворюються в самому організмі, називаються замінними. Амінокислоти, що не синтезуються в організмі людини, називаються незамінними. Вони повинні регулярно надходити з їжею. Білки, до складу яких входять замінні і незамінні амінокислоти в співвідношеннях, що наближаються до таких в організмі, називають повноцінними. Серед харчових продуктів практично немає білків, що цілком відповідають цим вимогам. Більш близькі до повноцінного білки материнського молока, курячого яйця. Отже, для повного забезпечення здорового організму повноцінними білками в добовий раціон повинні бути включені різні харчові продукти як тваринного, так і рослинного походження.
Для нормальної життєдіяльності людини необхідне надходження такої кількості повноцінного білка, що буде покривати всі потреби організму. Вони залежать від статі, віку, інтенсивності праці і т.д. З урахуванням цих факторів розроблені норми білкового харчування. Недостатнє споживання білків приводить до порушення процесів життєдіяльності, погіршення здоров'я, а тривале білкове голодування неминуче закінчується загибеллю.
Білки, необхідні для організму, насамперед, як пластичний матеріал, з якого будуються клітини всіх органів, тканин і систем. Однак харчові білки не можуть бути використані без попереднього розщеплення в організмі, тому що вони мають складну структуру і видову специфічність.
Розщеплення (гідроліз) білків на амінокислоти, що позбавлені видової і тканинної специфічності, відбувається в шлунково-кишковому тракті.
ПЕРЕВАРЮВАННЯ БІЛКІВ У ТРАВНОМУ ТРАКТІ. Перетравлення поживних речовин (білків, вуглеводів, ліпідів) – це процес гідролізу відповід-них сполук, що входять до складу продуктів харчування, відбувається в травно-му каналі і призводить до утворення простих біомолекул, які за рахунок дії спе-ціальних механізмів мембранного транспорту всмоктуються в кров або лімфу.
Переварювання білків починається в шлунку під дією шлункового соку. У його склад входить хлоридна кислота, що виробляється обкладовими клітками слизової оболонки шлунка. Вона денатурує білок, що полегшує його наступне розщеплення. До складу шлункового соку входять кислі фосфати та деякі органічні кислоти. Хлоридна кислота сприяє перетворенню проферменту пепсиногену, який секретується головними клітками слизової оболонки шлунка, в активний протеолітичний фермент пепсин.
Оптимальна концентрація водневих іонів для пепсину складає 1,5…2,5, що відповідає кислотності шлункового соку в процесі травлення. При збільшенні рН середовища до 6,0 (у кишечнику) пепсин втрачає свою активність. Пепсин відноситься до однокомпонентних ферментів, тобто до ферментів-протеїнів. За добу в шлунку виробляється близько 2 г пепсину. Каталітична активність пепсину шлунка дуже висока. Він каталізує розщеплення пептидних зв'язків у молекулі білка, утворених аміногрупами ароматичних і дикарбонових амінокислот. У результаті дії пепсину утворюються поліпептиди різної величини й окремі вільні амінокислоти.
Крім пепсину, у шлунковому соку міститься протеолітичний фермент гастріксин, оптимальні значення рН якого знаходяться в межах 3,5…4,5. Гастріксин вступає в дію на останніх етапах переварювання їжі в шлунку.
У шлунку грудних дітей виявлений сичуговий фермент – хімозин. Оптимум дії цього ферменту відповідає рН 3,5…4,0. Під впливом хімозину в присутності солей кальцію казеїноген молока в ході гідролізу перетворюється в казеїн і молоко згортається.
Легше інших у шлунку переварюються альбуміни і глобуліни тваринного і рослинного походження; погано розщеплюються білки сполучної тканини (колаген і еластин) і зовсім не розщеплюються кератин і протаміни.
Частково перетравлена напіврідка маса поживних сполук, що утворюється в шлунку (хімус) періодично надходить через пілоричний клапан у дванадцятипалу кишку. В цю ж частину травного каналу надходять із підшлункової залози протеолітичні ферменти та пептидази, які діють на пептиди, що надходять із шлунка. Каталітична дія цих ферментів відбувається в слабколужному середовищі (рН 7,5…8,0), яке утворюється наявними в кишковому соку бікарбонатами. Більшість ферментів протеолітичної дії, що функціонують у тонкій кишці, синтезуються в екзокринних клітинах підшлункової залози у вигляді проферментів, які активуються після їх надходження в дванадцятипалу кишку (трипсиноген, хімотрипсиноген, проеластаза, прокарбоксипептидази А і Б). Гідроліз білків та пептидів, що надходять із шлунка, відбувається як у порожнині тонкої кишки, так і на поверхні ентероцитів. Гідроліз білків та пептидів, що надходить із шлунка, відбувається як у порожнині тонкої кишки, так і на поверхні ентероцитів – пристінкове або мембранне травлення.
Сік підшлункової залози надходить у дванадцятипалу кишку і змішується з кишковим соком. Ця суміш містить протеолітичні ферменти, що розщеплюють білки, альбумози і пептони до невеликих пептидів, а потім до амінокислот. До протеолітичних ферментів відносяться трипсин, хімотрипсин, карбоксипептидази, амінопептидази і велика група три- і дипептидаз.
Трипсин знаходиться в соку підшлункової залози в неактивній формі, у виді проферменту трипсиногену. Його активація відбувається під дією ферменту кишкового соку – ентерокінази. Для процесу активування необхідні іони Са2+. Процес перетворення трипсиногену в трипсин здійснюється шляхом відщіплення невеликого пептиду з N-кінця пептидного ланцюга ферменту.
Трипсин гідролізує як нерозщеплені в шлунку білки, так і високомолекулярні пептиди, діючи головним чином на пептидні зв'язки між аргініном і лізином. Оптимум рН для трипсину складає 7,0…8,0. Трипсин робить порівняно неглибокий гідроліз білка, утворює поліпептиди і невелику кількість вільних амінокислот.
Активність трипсину може знижатися під впливом ряду інгібіторів. До них відносяться основні пептиди з молекулярною масою 9000 Да. Вони виявлені в підшлунковій залозі, крові, легенях, у бобах сої. Знижує активність трипсину і мукопротеїн, що міститься в сирих яйцях – авідин.
Хімотрипсин – другий протеолітичний фермент підшлункової залози. Він також секретується в неактивній формі, у виді хімотрипсиногену. Під дією трипсину хімотрипсиноген переходить в активний фермент – хімотрипсин. Дія хімотрипсину подібна дії трипсину. Оптимум рН для обох ферментів приблизно однаковий, хімотрипсин діє на білки і поліпептиди, що містять ароматичні амінокислоти (тирозин, фенілаланін, триптофан), а також на пептидні зв'язки, що не піддаються впливові трипсину (метіонін, лейцин).
Пептиди, що утворилися в результаті дії на білки пепсину, трипсину і хімотрипсину в нижніх відділах тонкої кишки, піддаються подальшому розщепленню. Цей процес здійснюють карбоксипептидази, амінопептидази. Ці ферменти відносяться до металоферментів. Вони активуються двовалентними іонами Mg2+, Mn2+, Со2+, що відіграють важливу роль у формуванні фермент-субстратного комплексу.
Механізм дії аміно- і карбоксипептидаз полягає у відщіпленні від пептидів кінцевих амінокислот, що мають вільну амінну або карбоксильну групу. Невеликі пептиди, що залишилися нерозщепленими і складаються з трьох-чотирьох амінокислотних залишків, піддаються гідролізу специфічними ди- і триамінопептидазами. Усоці підшлункової залози є фермент еластаза.
Еластаза – ендопептидаза, що також має широку субстратну специфічність, розщеплюючи пептидні звязки, що утворюються залишками амінокислот малого розміру – гліцину, аланіну, серину.
Таким чином, у результаті послідовної дії на білки протеолітичних ферментів у кишечнику утворюються вільні амінокислоти, що всмоктуються в кров через стінку кишечнику.
УТВОРЕННЯ В КИШЕЧНИКУ ОТРУТНИХ ПРОДУКТІВ РОЗПАДУ БІЛКІВ Й ЇХНЄ ЗНЕШКОДЖЕННЯ. Амінокислоти, що не всмокталися в кров через слизову оболонку тонкої кишки, піддаються впливу мікроорганізмів у товстому кишечнику. При цьому ферменти мікроорганізмів розщеплюють амінокислоти і перетворюють їх в аміни, жирні кислоти, спирти, феноли й інші речовини, нерідко отруйні для організму. Цей процес іноді називають гниттям білків у кишечнику. У його основі лежить декарбоксилювання амінокислот, при цьому з амінокислот з'являються біологічні аміни. Так, з амінокислоти орнітину утворюється путресцин:
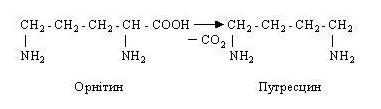
З лізину утворюється кадаверин:
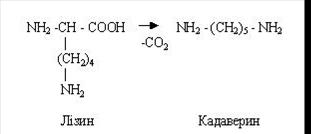
Путресцин і кадаверин виводяться з організму з фекальними масами. У тих випадках, коли ці сполуки попадають у кров, вони виводяться із сечею в незміненому виді.
З тирозину утворюється крезол, а якщо процес йде далі, то і фенол:
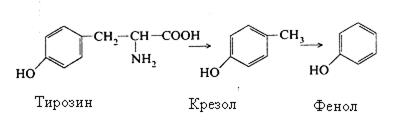
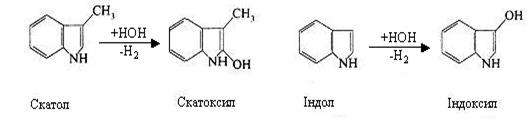
З триптофану утворюються скатол і індол:
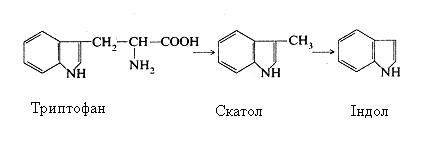
При глибокому руйнуванні кишковими мікроорганізмами сірковмісних амінокислот – цистіну, цистеїну і метіоніну – утворюється сірководень (H2S), меркаптан (CH3SH) і інші сірковмісні сполуки.
Продукти гниття білків всмоктуються у венозну кров, потім попадають у печінку, де і знешкоджуються за допомогою сірчаної або глюкуронової кислоти.
Цей процес здійснюється за наступною схемою:
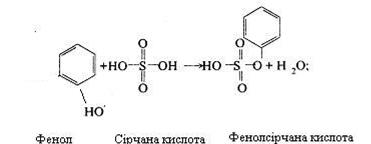
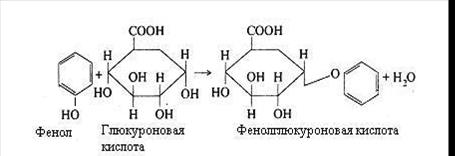
Індол і скатол також знешкоджуються в печінці при участі сірчаної і глюкуронової кислот. Однак вони попередньо окислюються: скатол у скатоксил, індол у індоксил і у виді парних кислот виводяться з організму із сечею.
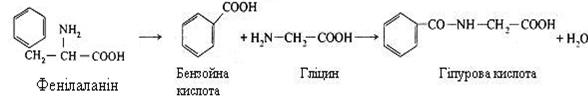
Деякі отруйні речовини, наприклад, бензойна кислота, що утворилася з фенілаланіна, знешкоджуються в печінці за допомогою гліцину. При цьому утворюється гіпурова кислота – нешкідлива сполука, що виділяється із сечею.
Можливості печінки в знешкодженні утворених в товстій кишці отруйних речовин, що всмокталися в кров, не безмежні. При зниженні її функціональної здатності (наприклад, у зв'язку з перенесеними раніше захворюваннями) надходження значної кількості отруйних речовин може виявитися надмірним навантаженням, тоді частина незнешкоджених отруйних речовин розноситься (великим колом кровообігу) по всьому організму, викликаючи його отруєння. Відбувається передчасне старіння клітин і їхня загибель. При цьому відзначається погіршення самопочуття людини, її мучать головні болі.
Для попередження негативного впливу отруйних речовин на організм необхідно раціонально планувати харчовий раціон. У нього повинні бути включені продукти, що містять не тільки білки, але і жири і вуглеводи, корисні кисломолочні продукти, тому що молочнокислі бактерії сприяють прискоренню загибелі гнильних мікроорганізмів товстої кишки. У раціоні необхідна їжа, що є джерелом пектинових речовин і клітковини, що, підвищуючи рухову активність кишечнику, сприяють виведенню шлаків (у тому числі й отруйних речовин) з організму.
обмін білків в тканинах. Основна частина амінокислот, що утворюється в кишечнику з білків, надходить у кров (95 %) і невелика частина – у лімфу. По ворітній вені амінокислоти попадають у печінку, де витрачаються для біосинтезу різних специфічних білків (альбумінів, глобулінів, фібриногену), інші амінокислоти струмом крові розносяться до всіх органів і тканин, транспортуються усередину кліток, де вони використовуються для біосинтезу білків. Невикористані амінокислоти окиснюються до кінцевих продуктів обміну.
Процес розщеплення тканинних білків каталізується тканинними ферментами - протеїназами-катепсинами.
Співвідношення між амінокислотами в білках, що розпадаються і синтезуються, різне, тому частина вільних амінокислот повинна бути перетворена в інші амінокислоти або окиснена до простих сполук і виведена з організму. Отже, в організмі існує внутрішньоклітинний запас амінокислот, що значною мірою поповнюється за рахунок процесів взаємоперетворення амінокислот, гідролізу білків, синтезу амінокислот і надходження їх з позаклітинної рідини. У той же час завдяки синтезові білків і інших реакцій (утворенню сечовини, пуринів і т.п.) постійно відбувається видалення вільних амінокислот з позаклітинної рідини.
Шляхи обміну амінокислот. В основі різних шляхів обміну амінокислот лежить три типи реакцій: за амінною і карбоксильною групами і за бічним ланцюгом. Реакції за амінною групою включають процеси дезамінування, переамінування, амінування, за карбоксильною групою – декарбоксилювання. Безазотиста частина карбогенного кістяка амінокислот піддається різним перетворенням з утворенням сполук, що потім можуть включатися в цикл Кребса для подальшого окиснювання.
Шляхи внутрішньоклітинного перетворення амінокислот складні і перехрещуються з багатьма іншими реакціями обміну, у результаті чого проміжні продукти обміну амінокислот можуть служити необхідним попередником для синтезу різних компонентів клітин і бути біологічно активними речовинами.
Катаболізм амінокислот у ссавців відбувається, в основному, в печінці і трохи слабкіше в нирках.
Дезамінування амінокислот. Суть дезамінування полягає в розщепленні амінокислот під дією ферментів на аміак і безазотистий залишок (жирні кислоти, оксикислоти, кетокислоти). Дезамінування може йти у виді відбудовного, гідролітичного, окисного і внутрімолекулярного процесів. Останні два типи переважають у людини і тварин.
Окисне дезамінування підрозділяється на дві стадії. Перша стадія є ферментативною, вона закінчується утворенням нестійкого проміжного продукту – імінокислоти, що у другій стадії спонтанно в присутності води розпадається на аміак і a-кетокислоту. Ферменти, які каталізують цей процес, містять у якості простетичної групи НАД або ФАД.
В організмі людини найбільше активно протікає дезамінування глутамінової кислоти під дією ферменту глутаматдегідрогенази, що знахо-диться в мітохондріях клітин усіх тканин. У результаті цього процесу утворює a-кетоглутарова кислота, що бере участь у багатьох процесах обміну речовин.
Трансамінування (переамінування) амінокислот. Обов'язковою умовою трансамінування є участь дикарбонових амінокислот (глутамінової і аспарагінової), що у виді відповідних їм кетокислот – a-кетоглутарової і щавелевооцтової можуть взаємодіяти з всіма амінокислотами, за винятком лізину, треоніну й аргініну.
При переамінуванні відбувається безпосередній перенос аміногрупи з амінокислоти на кетокислоту, а кетогрупи – з кетокислоти на амінокислоту без звільнення при цьому аміаку.
Цей процес протікає в кілька етапів. У загальному виді реакція виглядає так:
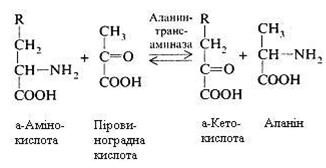
Реакцію каталізують ферменти, що відносяться до класу трансфераз, їх простетичною групою є фосфорпіридоксаль – фосфорний ефір вітаміну В6.
Процес переамінування широко розповсюджений у живій природі. Його особливість – легка оборотність.
Реакції переамінування відіграють велику роль в обміні речовин. Від них залежать такі найважливіші процеси, як біосинтез багатьох замінних амінокислот з відповідних їм кетокислот, розпад амінокислот, об'єднання шляхів вуглеводного й амінокислотного обміну, коли з продуктів розпаду глюкози, наприклад, піровиноградної кислоти, може утворитися амінокислота аланін і навпаки.
Відбудовне амінування. Цей процес протилежний дезамінуванню. Він забезпечує зв'язування аміаку кетокислотами з утворенням відповідних амінокислот.
Відбудовне амінування каталізується добре функціонуючою ферментною системою, що забезпечує амінування a-кетоглутарової або щавелевооцтової кислоти з утворенням глутамінової або аспарагінової кислоти.
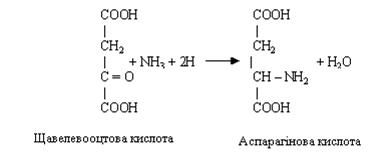
При знешкодженні аміаку неорганічними й органічними кислотами відбувається утворення амонійних солей. Цей процес здійснюється в нирках. Амонійні солі, що утворилися, виводяться з організму із сечею і потім.
Декарбоксилювання амінокислот. Процес декарбоксилювання каталі-зується декарбоксилазами, специфічними для кожної амінокислоти, простетичною групою яких служить піридоксальфосфат. Ці ферменти відносяться до класу ліаз. Процес декарбоксилювання, що полягає у відщіпленні від амінокислот СО2 з утворенням амінів, можна показати на наступній схемі:
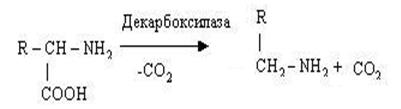
Механізм реакції декарбоксилювання амінокислот відповідно до загальної теорії піридоксалевого каталізу зводиться до утворення піридоксальфосфат-субстратного комплексу в активному центрі ферменту.
Таким шляхом із триптофану утворюється триптамін, з гідрокситриптофана – серотонін. З амінокислоти гістидину утворюється гістамін. З глутамінової кислоти при декарбоксилюванні утворюється g-аміномасляна кислота (ГАМК).
Аміни, утворені з амінокислот, називають біогенними амінами, тому що вони виявляють на організм могутній біологічний ефект.
Біогенні аміни виявляють фізіологічну дію в дуже малих концентраціях. Так, введення в організм гістамину приводить до розширення капілярів і підвищення їхньої проникності, звуження великих судин, скорочення гладких м'язів різних органів і тканин, підвищення секреції хлоридної кислоти в шлунку. Крім того, гістамін бере участь у передачі нервового порушення.
Серотонін сприяє підвищенню кров'яного тиску і звуженню бронхів; його малі дози придушують активність центральної нервової системи, у великих дозах ця речовина робить стимулюючу дію. У різних тканинах організму великі кількості гістаміну і серотоніну знаходяться в зв'язаній, неактивній формі. Біологічну дію вони виявляють тільки у вільній формі.
g-аміномасляна кислота (ГАМК) накопичується в мозковій тканині і являє собою нейрогуморальний інгібітор-медіатор гальмування центральної нервової системи.
Великі концентрації цих сполук можуть являти загрозу для нормального функціонування організму. Однак у тваринних тканинах мається аміноксидаза, що розщеплює аміни до відповідних альдегідів, що потім перетворюються в жирні кислоти і розпадаються до кінцевих продуктів.
ПРОЦЕСИ ЗНЕШКОДЖЕННЯ АМІАКУ. У процесі перетворення амінокислот у тканинах утворюються їхні кінцеві продукти обміну – оксид карбогену, вода й аміак. Вода використовується організмом для забезпечення біохімічних процесів. Оксид карбогену частково виводиться з організму з видихуваним повітрям, інша його частина утилізується в процесах синтезу (наприклад, при синтезі жирних кислот, пуринових основ і т.д.). Аміак, що утворюється в результаті дезамінування амінокислот, є токсичною речовиною, збільшення його концентрації в крові й інших тканинах робить несприятливу дію, особливо на нервову систему. Токсичність аміаку обумовлена тим, що він сприяє відбудовному амінуванню a-кетоглутарової кислоти в мітохондріях. Це приводить до видалення її з циклу Кребса і, як наслідок, до падіння тканинного дихання і надлишкового утворення кетонових тіл з ацетил-КоА.
У процесі еволюції живі організми виробили різні ефективні механізми зі знешкодження токсичної дії аміаку, основними з яких є: утворення амінів глутаміну або аспарагіну, відбудовне амінування, нейтралізація кислот, синтез сечовини.
Синтез глютаміну або аспарагіну має велике значення для організму. Він протікає в місцях безпосереднього утворення аміаку (наприклад, у печінці, мозку), там же знаходиться і фермент, каталізуючий цей процес, глутамінсинтетаза, що відноситься до класу лігаз. Синтез амідів вимагає доставки енергії у виді АТФ, присутності глутамінової або аспарагінової кислот, вільного аміаку і відповідних специфічних ферментів. Реакція синтезу аміду пов’язана з розпадом АТФ.
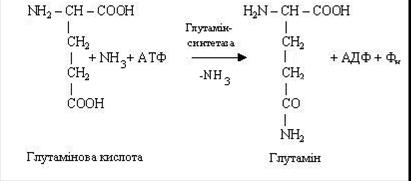
Аналогічно утворюється і аспарагін.
У результаті взаємодії аміаку з глутаміновою і аспарагіновою кислотами відбувається його зв'язування і в такий спосіб аміак знешкоджується. Зв'язаний аміак може бути використаний як джерело азоту (наприклад, для синтезу пуринових і піримідинових основ, мукополисахаридів). Глутамін і аспарагін не тільки знешкоджують аміак, але і виступають у якості його транспортної форми. У зв'язаному виді аміак доставляється до місця остаточної утилізації – у печінку, де з нього синтезується сечовина.
Синтез сечовини. Одним з найбільш ефективних методів знешкодження аміаку є синтез сечовини. Вперше схема синтезу сечовини була запропонована російським біохіміком М. В. Ненцьким, який вважав, що сечовина утворюється з двох молекул аміаку й однієї молекули вугільної кислоти, але ця теорія виявилася помилковою.
У спеціальних дослідах М.В. Залесського і С.С. Салазкіна на тваринах (лабораторія акад. І.П. Павлова) було встановлено, що якщо венозну кров направити не в печінку, а, минаючи її, у нижню порожню вену, то спостерігається різке збільшення вмісту аміаку в крові й отруєння. На підставі цих результатів був зроблений висновок, що печінка є органом, у якому відбувається знешкодження аміаку.
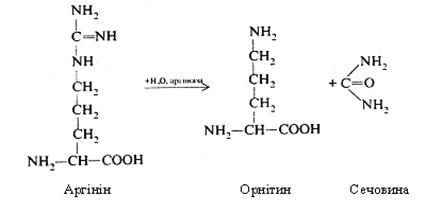
У роботах X.А. Кребса й інших дослідників була показана участь у даному процесі речовин і ферментів, каталізуючих реакції синтезу сечовини. Англійський біохімік Х. Кребс вніс великий вклад у сучасну теорію синтезу сечовини. Він встановив, що цей процес носить циклічний характер, і вказав на роль у ньому орнітину. Передумовою для створення теорії синтезу сечовини було виявлення в печінці амінокислоти орнітину і ферменту аргінази, що розщеплює аргінін на орнітин і сечовину. За теорії Кребса, синтез сечовини починається взаємодією орнітину з аміаком і оксидом карбогену. При цьому утворюється цитрулін, що взаємодіє з ще однією молекулою аміаку з виділенням аргініну. Останній гідролізується аргіназою на орнітин і сечовину. Орнітин у цих реакціях виконує роль каталізатора. За відкриття цього циклу X. А. Кребс був визнаний гідним Нобелівської премії.
Замкнутий цикл синтезу сечовини умовно можна розділити на три етапи. На першому і другому етапах відбувається зв'язування двох молекул аміаку в нешкідливу для організму сполуку, на третьому етапі утворюється сечовина.
Перший етап. За рахунок енергії АТФ із молекули аміаку й оксиду карбогену синтезується карбомоїлфосфат. Потім відбувається конденсація карбомоїлфосфату з орнітином, у результаті якої синтезується цитрулін. При цьому вивільняється неорганічний фосфат.
Синтез цитруліну забезпечується за рахунок енергії, акумульованої в карбомоїлфосфаті.
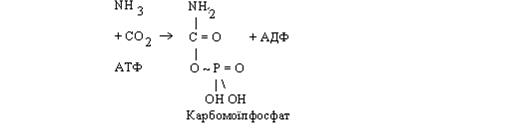
Таким чином, у цитруліні зафіксована одна молекула аміаку.
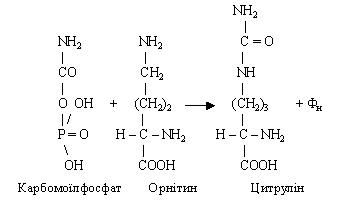
Другий етап. Відбувається зв'язування ще однієї молекули аміаку в глутамінову кислоту шляхом відбудовного амінування з a-кетоглутаровою кислотою. Глутамінова кислота передає зафіксовану молекулу аміаку у виді NH2-групи на щавелевооцтову кислоту, що перетворюється в аспарагінову кислоту (процес переамінування):
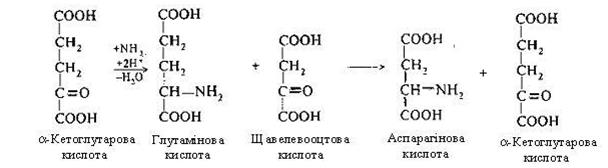
Третій етап. На цьому етапі здійснюється синтез сечовини. Цитрулін, взаємодіючи з аміногрупою аспарагінової кислоти, утворює аргінінобурштинову кислоту (аргінінсукцинат).
У цьому процесі використовується енергія ще однієї молекули АТФ.
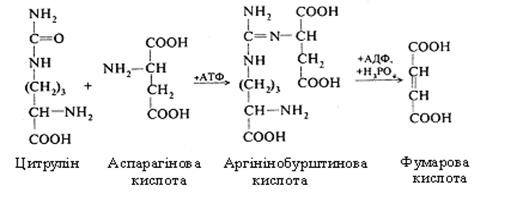
Аргінінобурштинова кислота розщеплюється на аргінін і фумарову кислоту. Фумарова кислота включається у вуглеводний обмін, перетворюючись в яблучну, котра шляхом дегідрування переходить у щавелевоцтову.
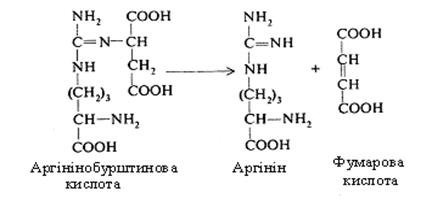
Щавелевооцтова кислота може приєднувати до себе аміак і знову перетворюватися в аспарагінову кислоту або, піддаючись декарбоксилюванню, у піровиноградну, а потім у коензим А, що використовується в різних реакціях біосинтезу або окисляється до СО2 і Н2О.
Аргінін, що утворився, під дією аргінази гідролітично розщеплюється на орнітин і сечовину. Орнітин, що вивільнився, може знову вступити в реакцію з новою молекулою карбомоїлфосфату і процес повториться.
Підраховано, що в стані азотистої рівноваги організм дорослої людини споживає і відповідно виділяє 15 г азоту; з екскретованого із сечею азоту на долю сечовини приходиться близько 85 %, креатініну – 5, амонійних солей – 3, сечової кислоти – 1, інших його форм – 6.
ОБМІН НУКЛЕЇНОВИХ КИСЛОТ В ОРГАНІЗМІ. Нуклеїнові кислоти входять до складу клітин у виді складних білків – нуклеопротеїдів, що надходять в організм із продуктами харчування. У шлунку під дією хлоридної кислоти і пепсину вони розщеплюються на прості білки і нуклеїнові кислоти. Хлоридна кислота розриває зв'язки між нуклеїновою кислотою і білками (гістаміни і протаміни), а інші, більш стійкі зв'язки при рН шлункового соку (1,5…2,5), розщеплює, головним чином, пепсин.
У тонкому кишечнику під дією трипсину, хімотрипсину і пептидаз білки розщеплюються до амінокислот, які всмоктуються слизовою оболонкою і надходять у кров. Нуклеїнові кислоти під впливом ферментів підшлункової залози і тонкого кишечнику – нуклеаз – розпадаються до мононуклеотидів. Останні під дією ферментів кишкового соку втрачають фосфорну кислоту і перетворюються в нуклеозиди. Далі нуклеозиди під дією нуклеозидаз розпадаються на складові частини – азотисті основи і пентози. Однак нуклеозидази тонкого кишечнику недостатньо активні і не можуть забезпечити розщеплення всієї маси нуклеотидів. У зв'язку з цим у кров всмоктуються в основному мононуклеотиди і нуклеозиди, що піддаються специфічним перетворенням у тканинах.
Тканинні нуклеїнові кислоти під дією нуклеаз розщеплюються до мононуклеотидів, а потім до азотистих основ, пентоз і фосфорної кислоти.
Фосфорна кислота бере участь у фосфорилюванні, а також у буферних системах, синтезі фосфоліпідів, фосфопротеїнів, АТФ і інших сполук. З організ-му виводиться переважно у виді кислих солей натрію разом із сечею. Пентози можуть окислятися до СО2 і Н2О або використовуватися для синтезу глюкози, нуклеотидів
Дата добавления: 2020-03-17; просмотров: 1348;
Поиск по сайту
Узнать еще
- II. ТЕОРИЯ ТЕПЛООБМІНУ
- Білковий обмін в організмі, що розвивається
- Вікова динаміка основного обміну
- Газообмін у легенях
- Енергетичний обмін – це метаболічні процеси, пов’язані з утворенням АТФ.
- Йонообмінна хроматографія
- Класифікація і основні схеми теплообмінних апаратів
- Лекція № 10. Обмін ліпідів в організмі.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
