Ассоциативное научение
Для рассмотрения гипотезы о том, что животные обладают механизмами обнаружения и узнавания причинных отношений, следует дать определение природы таких отношений. Существуют два основных типа причинных связей, и вряд ли можно сомневаться в том, что оба их животное может научиться различать (Dickinson, 1980). Одно событие (причина) может вызвать другое событие (следствие) или не вызвать его (отсутствие следствия). Первое событие не обязательно должно быть непосредственной причиной следствия или его отсутствия, но оно может быть определенным звеном причинно-следственной цепи. В самом деле, событие, замеченное животным, может быть не частью причинной цепи, а просто указанием на то, что произошло вызванное данной причиной событие.
Именно кажущаяся причина важна для животного.
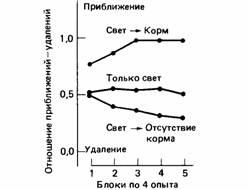
Способность научиться узнаванию обоих типов причинных отношений можно показать в простом опыте. Голодных голубей сажают в скиннеровский ящик с двумя световыми ключами и механизмом подачи корма. Одной группе голубей демонстрируют связь свет-корм (причина-следствие), а другой свет - отсутствие корма (причина без следствия). Третьей группе предъявляют только свет. В первом случае один из ключевых дисков освещается на 10 с с неправильными интервалами и пища подается, как только выключается свет. Во втором случае свет и пищу предъявляют столько же раз, что и в первом, но внимательно следят за тем, чтобы корм никогда не появлялся вслед за включением света. В третьем случае совсем не дают пищи. В первом случае свет сигнализирует о появлении корма, а во втором - о его отсутствии. Затем свет можно предъявить каждой группе голубей отдельно от пищи и отметить, приблизится к нему птица или, наоборот, удалится. Результаты такого опыта (рис. 19.5) ясно показывают, что голуби, наблюдавшие связь свет корм, стремятся приблизиться к свету, как и можно ожидать на основе нормальной классической выработки условного рефлекса. Голуби, которым предъявлялось отношение свет-отсутствие корма, также не оставались безразличными к свету, а определенно избегали его (Wasserman et al., 1974). Лишь голуби третьей группы были, по-видимому, безразличны к свету (рис. 19.5).
Анализируя результаты этого опыта, надо помнить о ряде важных моментов. Во-первых, мы не можем считать, что голубь научился устанавливать причинную связь света с пищей или ее отсутствием. То, чему животные научаются, сталкиваясь с причинными отношениями, можно рассматривать только с позиций теорий о внутренних изменениях, происходящих в результате приобретенного опыта (см. ниже). Во-вторых, тот факт, что в одном случае голуби приближаются к свету, а в другом удаляются от него, интересен, но не относится непосредственно к обсуждаемой проблеме. Многие раз-
|
| Рис. 19.5.Приобретение голубями, которым предъявляются разные сочетания освещения ключа и подачи корма, тенденции приближаться к освещенному ключу или отдаляться от него. В условиях «свет-корм» освещение ключа сочеталось с кормом и голуби вырабатывали реакцию приближения. В условиях применения только света голуби никогда не получали корма и никак не реагировали на ключ. В условиях «свет - отсутствие корма», они получали корм, но ни во время, ни вскоре после освещения ключа; у этих голубей возникала реакция удаления от ключа. (По Dickinson, 1980.) |
линия в поведении, вызванные предъявлением стимулов, можно было бы счесть указанием на произошедшее научение, однако говорить о значении конкретных действий животного логично только при анализе того, чему именно оно научилось. В-третьих, имеет смысл считать свет кажущейся причиной непредъявления пищи, только если пища предъявляется в его отсутствие. Нельзя ожидать, что голубь научится принимать свет за причину отсутствия пищи, если он никогда не получал ее в данной ситуации, как это было в третьей опытной группе. Животное имеет возможность усвоить ассоциацию «причина-отсутствие следствия», только когда почему-либо ожидает, что определенное следствие произойдет (Dickinson, 1980).
Хотя иногда можно сделать вывод, что изменение поведения говорит о произошедшем научении, отсутствие таких изменений нельзя считать указанием на полное отсутствие научения. Дикинсон (1980) называет это проблемой молчания поведения. Так, например, в случае описанного
выше опыта мы не можем считать, что голуби, которым предъявлялся только свет, ничему не научились. Напротив, имеются данные о том, что крысы научаются игнорировать стимулы, не предсказывающие никакого изменения в ответ на их действия (Mackintosh, 1973; Baker, Mackintosh, 1977). Если в ходе опыта крысам вначале предъявляют два стимула независимо один от другого, то потом они научаются связывать их медленнее, чем животные, которым эти стимулы предварительно не предъявлялись. Следовательно, крысы научаются тому, что определенные стимулы незначимы, и это мешает последующему основанному на этих сигналах научению.
Существуют и другие формы научения, при которых наблюдается молчание поведения. Животные могут научиться тому, что два события не связаны между собой, т. е. следствие не связано либо с данной причиной, либо с целым классом причинных событий. Если этот класс действий входит в поведенческий репертуар животного, то такую форму научения называют «приобретенной беспомощностью» (Maier, Seligman, 1976), т.е. животные научаются тому, что они ничего не могут сделать для улучшения ситуации. Такая выученная беспомощность замедляет будущее научение в аналогичных условиях.
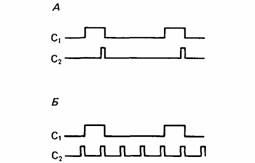
Для того чтобы животное научилось различать простую причинную связь, между двумя событиями должна быть полная положительная корреляция (Dickinson, 1980). С точки зрения животного, всегда имеется множество возможных причин события помимо той, которую применил экспериментатор. Как показано на рис. 19.6, между двумя событиями должна существовать достаточно тесная связь, для того чтобы одно из них было принято за причину второго. Для выяснения значения фоновых, или ситуационных, сигналов Макинтош (Mackintosh, 1976) обучал крыс нажимать на рычаг для получения корма, а затем вводил в опыт различные стимулы. Одной группе при каждом нажиме предъявляли световой сигнал, другим - сложный, состоявший из света и шума. Для одних групп исполь-
|
| Рис. 19.6. Распределение событий при разных соотношениях между событиями С1 и С2 . А. С2 происходит только во время или вскоре после С1 , и оба они коррелируют между собой положительно. Б. Появление С2 в равной мере вероятно в отсутствии или в присутствии С1 , и между ними нет корреляции. (По Dickinson, 1980.) |
зовался слабый шум (50 дБ), для других -сильный (85 дБ). Во всех случаях животные сразу после предъявления каждого стимула получали легкий удар током. В конце опыта всем группам предъявляли один только свет, чтобы узнать, насколько животные научились связывать его с током.
Полученные результаты (рис. 19.7) показывают, что свет подавлял нажимы на рычаг в разной степени в разных группах. Если предъявлялся только свет или свет со слабым шумом, подавление было значительным, а когда свет сопровождался сильным шумом, оно было гораздо слабее. Таким образом, наличие мощного второго стимула ослабляло связь света с током даже несмотря на то, что между ними была полная корреляция. Это явление называется затенением. Степень затенения зависит от сравнительной силы затеняемого и затеняющего стимулов, поэтому слабый шум производил незначительное затеняющее действие.
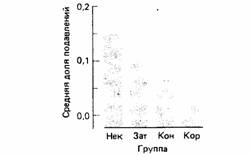
Животные научаются связывать два события, только если вначале сочетание их было неожиданным или внезапным (Mackintosh, 1974). В обычном опыте по условным рефлексам такая внезапность создается подкреплением. Так, если стимул сопровождается ударом тока, причем ни сам этот стимул, ни фоновые сигналы в начале эксперимента не предвещали включения тока, такое подкрепление будет внезапным. Но предположим, что животное уже испытало раньше удар тока в присутствии стимула А; тогда, если с током коррелируют стимулы А и Б, присутствие первого из них будет блокировать выработку реакции на второй. Это явление, впервые открытое Кеймином (Kamin, 1969), называют блокадой. В одном из своих экспериментов Рескориа (Rescoria, 1971) продемонстрировал этот эффект и показал также, что чем неожиданнее подкрепление, тем лучше животное научается (рис. 19.8).
В природе временная связь между какими-либо событиями не всегда является причинной связью. Например, если кошка прыгает на яблоню, когда лает собака, а
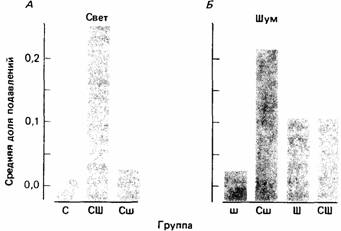
| Рис. 19.7. Степень подавления светом (А) или шумом (Б) нажимов на рычаг, производимых для получения корма крысами, которым предварительно предъявлялись сочетания удара током со светом (С), слабым шумом (ш), сильным шумом (Ш), светом вместе сослабым шумом (Сш) или светом вместе с сильным шумом (СШ). Большие числа означают меньшее подавление. (По Mackintosh, 1976.) |
|
| Рис. 19.8. Степень подавления стимулом А нажимов на рычаг, совершаемых для получения корма крысами, которым предварительно предъявлялись сочетания сложного стимула АБ с током. Некоррелирующей группе (Нек) предварительно предъявлялись сочетания Б с ударом тока. Группе с простым затенением (Зат) предварительно предъявлялся только ток. Контрольной группе (Кон) предварительно в случайном порядке предъявлялись Б и ток. Коррелирующей группе (Кор) предъявлялась отрицательная корреляция Б с током. Большие цифры означают меньшее подавление. (По Rescorla, 1971.) |
затем на землю падает яблоко, то мы скорее подумаем, что кошка, а не лай собаки, была причиной его падения. Как прыжок кошки, так и лай собаки одинаково связаны во времени с падением яблока, но другие детали этих событий приводят нас к заключению, что его причиной была все же кошка. Точно так же мы можем показать, что животные легче образуют ассоциации между одними типами стимулов, чем между другими. Например, крысы легко связывают вкус с последующей болезнью, но нелегко научаются связывать с ней тон или свет (Domjan, Wilson, 1972; см. также гл. 18). Показано, что крысы быстрее ассоциируют между собой два события, воспринимаемые одной сенсорной модальностью (Rescorla, Furrow, 1977) или локализованные в одном месте (Testa, 1975; Rescorla, Cunningham, 1979).
Наконец, для того чтобы животное связало между собой два события, обычно нужно, чтобы они были очень близки во времени. Временные отношения между двумя событиями изучались во многих опытах по научению (см. Dickinson, 1980), показавших, что оно наиболее эффективно, когда одно событие (причина) происходит незадолго до второго (следствия). Однако некоторые данные говорят в пользу того, что такие отношения объясняются не прямым действием временного интервала на процесс научения, а различной степенью затенения первого события фоновыми сигналами в зависимости от временного интервала (Dickinson, 1980). А это означает, что влияние временного интервала на научение должно зависеть от значимости фоновых сигналов. В частности, следует ожидать, что фоновые сигналы будут гораздо менее важны при выработке ассоциации вкуса с болезнью, чем при сочетании тона с током, потому что присутствующие в экспериментальной ситуации фоновые стимулы обычно не существенны для научения избеганию определенного вкуса.
Ревуски (Revusky, 1971) первый предположил, что отсутствием затенения, возможно, объясняется выработка вкусового отвращения, которая происходит несмотря на очень длинные интервалы времени между едой и последующим заболеванием. Он показал, что эффективный интервал можно сократить, введя фон из значимых (вкусовых) стимулов.
Подводя итог, можно сказать, что животные научаются ассоциировать два события, если отношение между ними соответствует тому, что обычно называют причинно-следственной связью. Таким образом, они способны усвоить, что одно событие (причина) предвещает второе (следствие) или говорит о его отсутствии в будущем (в отсутствие следствия). Животные могут также научиться связывать некоторые стимулы с отсутствием последствий в данной ситуации или не придавать причинного значения определенному классу стимулов (включая собственное поведение животного). Условия для такого рода ассоциативного научения существуют, если исходить из гипотезы, что животные приспособлены для приобретения знаний о причинных связях в окружающей их среде. Итак, животное должно уметь отличать потенциальные причины от фоновых стимулов, а для этого должно произойти что-то неожиданное, что привлечет внимание животного к данным со-
бытиям, или же сами события должны быть (врожденно) значимыми в отношении определенных последствий. Если эти условия не выполняются, фоновые сигналы могут затенить потенциальные причинные события или же научение может быть блокировано предыдущей связью с незначимым в данный момент стимулом. Таким образом, условия ассоциативного научения соответствуют нашим опирающимся на здравый смысл взглядам на природу причинности. Они не согласуются с традиционной точкой зрения на научение как на автоматическую связь между стимулом и реакцией. Феномен усваивания информации при молчании поведения наводит на мысль о необходимости некой когнитивной интерпретации научения животного. Но это не должно приводить нас к поспешным выводам о когнитивных способностях животных или о природе их ума.
Репрезентации
Репрезентация обычно считается центральным вопросом при обсуждении мышления животных. Обладают ли животные внутренними репрезентациями, т. е. мысленными образами, отыскиваемых предметов или сложных пространственных или общественных ситуаций (Kummer, 1982)? Этот вопрос привлек значительное внимание философов (например, Dennett, 1978) и специалистов во многих отраслях науки о поведении.
Мы видели, что животные научаются связывать два события, если отношение между ними соответствует тому, что обычно называется причинной связью. Некоторые из условий, при которых происходит ассоциативное научение, не совпадают с традиционным воззрением на научение животного как на близкое к автоматическому образование связи между стимулом и реакцией. Они скорее соответствуют взглядам, согласно которым животные способны приобретать знания о причинных связях в окружающей среде.
Рассматривая природу внутренней репрезентации, кодирующей приобретенный опыт, Дикинсон (Dickinson, 1980) проводит различие между декларативной и процедурной репрезентациями. Декларативная репрезентация - это мысленный образ желаемого объекта или цели. Когда крыса использует декларативную репрезентацию, отыскивая корм в знакомом лабиринте, у нее имеется мысленный образ пищи и она знает, что должна выбрать, скажем, левый поворот, чтобы найти ее. Процедурная репрезентация - это совокупность команд, автоматически ведущих к желаемому объекту без формирования его образа. Таким образом, если крыса пользуется процедурной репрезентацией, чтобы найти пищу, она направляется в левый поворот не потому, что «знает» о находящейся там пище, а потому, что связывает левый поворот с ее получением (рис. 19.9).
В декларативной системе знание представлено в форме, соответствующей утверждению или предположению, описывающему отношение между событиями в окружающем животное мире, т.е. в форме репрезентации, которая не заставляет животное использовать информацию каким-либо особым способом. В процедурной системе форма репрезентации непосредственно отражает предстоящее применение знания. Так, например, Холланд (Holland, 1977) демонстрировал крысам сочетание тона с пищей, предъявляя время от времени тон длительностью восемь секунд, после чего в кормушке появлялся корм. Он заметил, что у крыс вырабатывается тенденция приближаться к кормушке при включении тона. Это наблюдение может говорить о том, что во время научения устанавливается порядок действий в отношении кормушки, и приобретенная информация откладывается в памяти в форме, тесно связанной с ее применением. Согласно альтернативному объяснению, крыса воспринимает тон как причину появления пищи, т.е. возникает декларативная репрезентация. То, что крыса стремится приблизиться к кормушке при включении тона, в таком случае должно объясняться иначе, потому что декларативная система пассивна, т. е. не управляет поведением животного. Таким образом, процедурная репрезентация дает менее сложное объяснение поведению крысы.
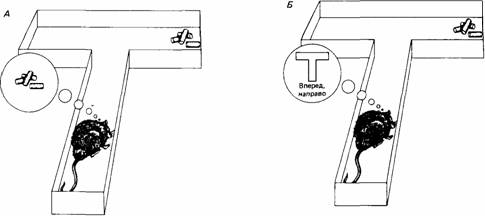
Рис. 19.9.Простые примеры декларативной (слева) и процедурной (справа) репрезентаций. В первом случае крыса обладает мысленным образом цели, во втором следует простому правилу поведения.
Предположим, однако, что после образования связи тон-пища крысы начинают сталкиваться с сочетанием пища-болезнь до тех пор, пока не станут отказываться от предлагаемого корма. Теперь у них образовались две отдельные ассоциации: тон-пища и пища-болезнь. Вопрос состоит в том, способны ли животные интегрировать их. С одной стороны, декларативная система дает основу для интеграции, потому что обе репрезентации имеют общий член «пища». Холланд и Штрауб (Holland, Straub, 1979) показали, что крысы интегрировали информацию, заключающуюся в двух таких связях, усвоенных в разное время. Животные, сталкиваясь с сочетанием пища-болезнь вслед за сочетанием тон-пища, были не склонны приближаться к кормушке при новом предъявлении тона.
Не приходится сомневаться в способности животных интегрировать возникшие по отдельности ассоциации, что легче всего объяснить декларативной системой. Однако при некоторых обстоятельствах интеграции не происходит, что говорит о возможности формирования поведения на основе процедурной репрезентации (Dickinson, 1980).
В декларативной системе должен быть какой-то способ перевода заложенной в памяти репрезентации во внешнее поведение. Для этого предложены различные механизмы (Dickinson, 1980), но здесь мы не будем на них останавливаться. Важно то, что, хотя процедурная теория дает относительно простое объяснение ситуаций простого научения, для истолкования наблюдаемых явлений может потребоваться более сложная теория, не исключающая некоторой формы декларативной репрезентации. Если допустить, что декларативная система необходима для объяснения поведения, необходимо также признать наличие у животного некоторой формы мышления. В то же время надо четко отделять свидетельства декларативной репрезентации от попыток упрощенного объяснения с помощью этого понятия поведенческих феноменов. Возможно, концепция декларативной системы - всего лишь удобный костыль, на который может опереться современная теория обучения.
ДЛЯ ЗАПОМИНАНИЯ
1. Некоторые стороны выработки условных рефлексов скрыты от наблюдателя, и это позволяет думать, что она включает когнитивные процессы.
2. О животных, которые на первый взгляд внезапно находят решение задачи, иногда говорят, что они проявили инсайт. Но не всегда ясно, чем именно такой процесс отличается от обыкновенного научения.
3. О некоторых сторонах ассоциативного научения говорят, что они требуют когнитивного объяснения, так как, по-видимому, связаны с мысленным образом достигаемой цели. Альтернативное объяснение предполагает, что животные просто выполняют сложный порядок действий.
Рекомендуемая литература
Dickinson A. (1980) Contemporary Animal Learning Theory, Cambridge University Press, Cambridge.
Часть III.
Дата добавления: 2016-06-18; просмотров: 3098;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
