Стратегия полов и социальная организация
Социальная организация у животных разных видов чрезвычайно различна: от простой кооперации между самцом и самкой до сложных сообществ, как у некоторых приматов. Одним из важнейших факторов, влияющих на социальную организацию вида, является система размножения данного вида. Чтобы понять разные системы размножения, нам надо оценить альтернативные эволюционные стратегии, возможные для самцов и самок.
Стратегия полов
В гл. 8. мы рассмотрели неизбежные различия в исходных вкладах самцов и самок в размножение. Самцы продуцируют огромное число мелких спермиев, а самки - относительно немного крупных яиц. Спермии испытывают сильную конкуренцию за возможность оплодотворить яйца, и самцы могут достичь наибольшего успеха в размножении, оплодотворяя много яиц при относительно небольших затратах на яйцо. Самка с самого начала вносит больший вклад в каждого потомка, и у большинства видов самки продолжают вкладывать больше, чем самцы, в доведение каждою детеныша до стадии, когда он способен выжить самостоятельно. Однако самка достигла бы более значительного успеха в размножении, если бы смогла заставить самца вносить больший вклад в выращивание их потомства, предоставив ей возможность отдать больше сил следующему поколению. А поскольку часто в интересах самца уменьшить свой вклад в воспитание потомства, между полами существует эволюционный конфликт.
Основная эволюционная проблема для самцов привлечь самок при наличии конкуренции со стороны других самцов. Основная эволюционная проблема для самок - выбрать в качестве полового партнера такого самца, который обеспечит потомство наилучшими шансами на выживание и размножение. По-видимому, самка с самого начала имеет преимущество, так как она может отказаться от спаривания. Как мы видели в гл. 8, обычно самка выбирает себе партнера во время брачных турниров. Самец немногого может добиться, если самка eго не принимает. Казалось бы, самец может заставить самку спариваться, однако обычно это довольно трудно, так как у большинства видов самка должна принять особую позу, чтобы копуляция стала возможна. Однако у некоторых животных, в том числе у крякв и скорпионовой мухи (Panorpa), иногда наблюдается принудительная копуляция. Самец скорпионовой мухи во время ухаживания обычно приносит самке свадебный подарок, часто это мертвое насекомое, вытащенное из тенет паука. Копуляция происходит, пока самка ест этот подарок. Иногда самец принуждает самку к копуляции и без подарка (Thornhill. 1980). В этом случае при успешной принудительной копуляции самец оказывается в выигрыше, поскольку ему не нужно рисковать во время поисков свадебного подарка. Известно, что 65% взрослых самцов скорпионовой мухи гибнет в паутине. Самка, не получая подарка, обычно обеспечивающего развивающиеся яйца энергией, оказывается в проигрыше. Однако степень успешности принудительного спаривания, очевидно, невелика, поэтому к такой стратегии насекомые прибегают в крайнем случае.
|
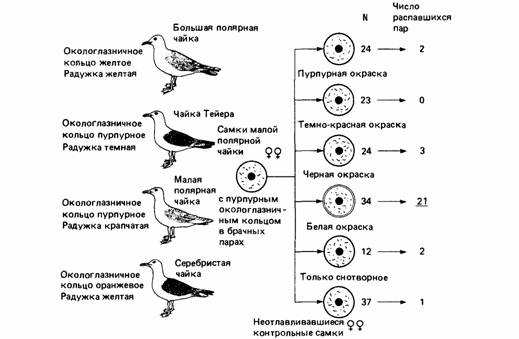
| Рис. 10.1. Изолирующие механизмы у чаек. Слева показаны внешние признаки, которые, как считают, играют важную роль для опознания друг друга особями одного вида. Результаты изменения цвета окологлазничного кольца у самок в брачных парах больших полярных чаек показаны справа. Числа в правой колонке показывают число пар, распавшихся в результате экспериментального вмешательства. Когда пурпурный цвет окологлазничного кольца был изменен на белый, 21 из 34 пар распались, тогда как в других экспериментах и в контроле распад брачных пар был невелик. (По Smith, I966.) |
Самцы обычно остаются в выигрыше в каждом сексуальном взаимодействии с самкой, если спариваются с ней, тогда как самки не всегда выигрывают и могут даже поплатиться снижением приспособленности (Parker, 1979). Поэтому для самок наилучшая стратегия быть скромной. Джордж Уильяме (Williams, 1966) описал ухаживание как спор между самцом, навязывающим товар, и самкой, не желающей его брать. Самке не надо было бы оставаться недоступной, если бы она прямо могла оценить приспособленность самца и его будущее поведение. Однако все, о чем самка может судить, это внешний вид самца и его поведение в настоящий момент. Самка должна заставить самца проявить свою истинную натуру или повернуть дело так, чтобы его интересы совпали с ее интересами. В обоих случах наилучший способ поведения для самки - быть осторожной и продлить период ухаживания. Первая задача для самки - удостовериться, что ее потенциальный партнер - самец того же вида. Это важно и для самца, однако, как мы уже видели, самка в случае ошибки теряет гораздо больше. Естественный отбор благоприятствует особям с отчетливыми признаками и поведением, позволяющими идентифицировать их как представителей конкретных видов. Совокупность этих признаков и поведения играет очень важную роль в ухаживании и называется изолирующими механизмами, поскольку эти механизмы обеспечивают репродуктивную изоляцию между видами, снижая возможность гибридизации. Обсудим вкратце три примера изолирующих механизмов.
1. На арктическом побережье Канады обитают четыре близкородственных вида чаек. В некоторых местах все четыре вида
сосуществуют в одной колонии, но при этом не скрещиваются друг с другом. Птицы чрезвычайно похожи и различаются только отметинами на концах крыльев и окраской окологлазничной области. Изменяя в эксперименте эти цветовые отметины, можно добиться спаривания разных видов (рис. 10.1) (Smith, 1966).
2. Обширные исследования скрещивания различных пород домашних кур показали, что внешний вид птицы очень важен (Lill, 1966, 1968) Так, например, во время ухаживания петух обходит курицу вальсирующими движениями и при этом демонстрирует оперение. При согласии спариваться куры припадают к земле в типичной позе готовности. После этого петух делает садку. Поскольку куры оценивают своих партнеров в основном по внешнему виду, а не по различиям в поведении, экспериментальное изменение различных типов окраски петухов (рис. 10.2) уменьшало их привлекательность для кур.
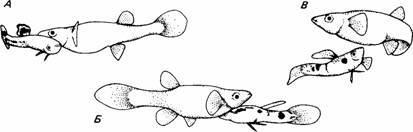
3. Известно, что самцы гуппи при ухаживании не делают различия в выборе самки, тогда как самки довольно строго выбирают партнера, основываясь на окраске самца и его поведении при ухаживании. Проводя эксперименты в аквариуме, Лили (Liley, 1966) обнаружил, что самцы видов Poecilia picta и Poecilia reticulata (рис. 10.3) часто ухаживают и делают попытки спариться с самками и своего, и чужого вида. А готовые к спариванию самки принимали ухаживания самцов только своего, но не другого вида. Однако, если у самок не было выбора, в конце концов они спаривались и с самцами другого вида.
Самка должна сделать все, чтобы удостовериться, что она спаривается с половозрелым и полноценным в половом отношении самцом. Продлевая период ухаживания, она с большей вероятностью будет спариваться с самцом, испытывающим сильную мотивацию и мощным в половом отношении. У европейского обыкновенного тритона (Triturus vulgaris) (рис. 10.4) интенсивность ухаживания самца зависит от числа сперматофоров (капсул со спермой), которые самец продуцирует во время данной встречи (Halliday,

| Рис. 10.2. Коричневый петух леггорн с прикрепленными белыми перьями, чтобы изменить цвет оперения на шее. (Из The Oxford Companion to Animal Behaviour, 1981.) |
1976). Во время ухаживания самец откладывает сперматофор на дно водоема; вероятность того, что самка успешно подберет его, возрастает в процессе ухаживания (Halliday, 1974). Чтобы отложить три сперматофора, самец должен оставаться под водой не дыша гораздо дольше обычного (Halliday, 1977a). Таким образом, самка, затягивая процедуру ухаживания, возможно, оценивает приспособленность самца.
Оценка самца самкой облегчается, если самец может продемонстрировать, что он являет собой достойный выбор. Некоторые исследователи полагают, что в этом отношении важную роль играет ритуальное кормление. Ритуальное кормление, во время которого самец подносит самке пищу или напоминающий пищу предмет, наблюдается у многих видов. У серебристой чайки (Lams argentatus) самка выпрашивает пищу, как если бы она была птенцом, а самец отрыгивает пищевой комок, который самка затем съедает. У большин-
|
| Рис. 10.3. А. Брачное поведение гуппи (Pvecilia picta). Самец кружится перед носом самки, демонстрируя яркую окраску хвостового и спинного плавников. Б. Сигмовидная поза у Poecilia reticulata. Самец принимает эту позу перед самкой и дрожит в течение нескольких секунд. Окраска самцов очень яркая и изменчивая. В. Попытка спариться у самца P. reticulata. После демонстрации окраски самец поворачивается кругом и пытается ввести гоноподий (модифицированный анальный плавник) в генитальное отверстие самки. Оплодотворение внутреннее. (Из The Oxford Companion to Animal Behaviour, 1981; по фотографиям Robin Liley.) |
ства видов количество пищи, которое самец передает самке, невелико по сравнению с ее обычным дневным рационом. У мухоловки-пеструшки (Ficedula hypoleuca), однако, это почти половина того, что обычно получает птенец такой же массы. У некоторых видов ритуальное кормление формализовано (см. гл. 22.1) и подношения реальной пищи не происходит. Ритуальное кормление влияет на вес кладки у обыкновенной крачки (Sterna hirundo) и может служить показателем будущего участия самца в выкармливании птенцов (Nisbet, 1973; 1977). Вирджиния Нибур (Niebuhr, 1981) установила, что ритуальное кормление у серебристой чайки было надежным показателем последующего участия самца в выкармливании птенцов. Оно также указывает на склонность самцов участвовать в насиживании и защищать птенцов после вылупления. По этим причинам ритуальное кормление - хороший показатель качеств самца как родителя.
Как и в случае с оценкой боевых способностей, здесь трудно найти критерий качеств самца, при демонстрации которых было бы невозможно смошенничать. Наилучшей стратегией самки в этом случае будет просто продлить или усложнить процесс ухаживания, так как это затруднит спаривание самца с другими самками, в особенности если сезон размножения короткий или мало свободных самок; иными словами, самцы, затрачивающие много сил и времени в процессе деятельности, предшествующей спариванию, такой, как ухаживание, строительство гнезда, защита территории, будут располагать меньшими временными и энергетическими ресурсами, чтобы преследовать других самок. Менее вероятно, что самец, уже затратив так много усилий на одну самку, оставит ее, вынуждая, таким образом, заботиться о потомстве в одиночестве. Более того, если самец утратит возможность спариваться с другими самками, он сможет увеличить свою приспособленность, только вкладывая оставшиеся ресурсы в свое потомство. Доукинс (Dawkins, 1976) назвал это «ситуацией семейного счастья».
Нет никаких сомнений, что «семейное счастье» можно наблюдать у некоторых моногамных видов. Однако это не является эволюционно стабильной ситуацией при всех обстоятельствах, так как здесь возможно вмешательство самок, придерживающихся иной стратегии. Доукинс создал модель, в которой возможны две стратегии для самок - «скромная» и «легкомысленная» и две для самцов - «верный» и «волокита». Скромные самки требуют продолжительного ухаживания перед копуляцией; легкомысленные самки копулируют без предварительного ухаживания. Верные самцы добровольно ведут длительное ухаживание, а после спаривания помогают самке вырастить потомство. Самцы-волокиты долго не ухаживают

|
| Рис. 10.4. Самец обыкновенного тритона (......) (Фотография ........ Halliday.) |
и, если не удастся спариться вскоре после встречи с самкой, оставляют ее и ищут другую. Самцы-волокиты могут оставить самку и после копуляции.
Теперь мы можем проанализировать эту ситуацию, чтобы решить, является ли это эволюцнонно стабильной стратегией (см. гл. 7). Пусть генетический выигрыш, получаемый каждым из родителей за одного выращенного потомка, будет оцениваться в +15 условных баллов. Затраты на выращивание одного потомка – 20 баллов, а затраты на продолжительное ухаживание –3 балла. В популяции, в которой все самки скромные и все самцы верные, размножающаяся пара поделила бы затраты на выращивание каждого потомка. Каждый из родителей получил бы + 15 баллов за каждого выращенного потомка. Поэтому за первого потомка результат в среднем был бы + 15 – 10 – 3 = +2. Хотя в реальной жизни родители могут и не нести затрат на ухаживание, допустим, что в рассматриваемой ситуации это происходит. Если теперь в популяции окажется легкомысленная самка, ей не придётся нести никаких затрат на ухаживание. При спаривании с верным самцом ее средний результат будет + 15 – 10 = + 5. что лучше результата скромной самки. Поэтому гены легкомысленного поведения будут распространяться среди самок популяции.
В популяции, где доля легкомысленных самок высока, возможно распространение и самцов-волокит. Они не добьются успеха при попытках спариться со скромными самками, но поскольку они быстро оставляют самок, которые не спариваются немедленно, волокиты немного теряют от таких встреч. А спариваясь с легкомысленной самкой, волокиты преуспевают. Они получают + 15 баллов за каждого выросшего потомка, но они ничего не тратят ни на ухаживание, ни на заботу о потомстве. Партнер волокиты, - легкомысленная самка, - вынуждена нести все — 20 баллов затрат при выращивании потомства. В результате спаривания с волокитой ее средний результат будет + 15 – 20 = – 5. Эта модель стабильна в том, что она не эволюционирует к крайнему состоянию, но она не стабильна в том, что не устанавливает конкретное сочетание стратегий самцов и самок; вместо этого она колеблется между альтернатив-
ными сочетаниями. Основываясь на эволюционной теории, можно было бы ожидать, что альтернативные стратегии самок обнаруживаются среди особей одного вида, как предсказывает модель Доукинса (Dawkins, 1976), однако оказывается, что они распространены не очень широко. Возможно, эти модели слишком просты или в них нет основных составляющих, о существовании которых мы и не подозреваем. Однако известно несколько примеров альтернативных стратегий самок.
У африканских страусов одни самки насиживают яйца, а другие не насиживают. Самец страуса удерживает большую территорию и строит что-то вроде гнезда на земле. Он спаривается с самкой, которая начинает откладывать в это гнездо яйца. В течение нескольких дней другие самки, которые могут спариваться, а могут и не спариваться с хозяином гнезда, начинают откладывать яйца в то же самое гнездо. Однако они не насиживают и не охраняют яйца, оставляя их на самца или его супругу. В конечном счете в гнезде может оказаться от 30 до 40 яиц, что слишком много для насиживания одной птицей. Насиживающая самка отодвигает некоторые яйца к краю гнезда, и они обычно гибнут, перегревшись на солнце, или их утаскивают хищники.
Бертран (Bertran, 1979) обнаружил, что самка страуса узнает собственные яйца и старается вытолкнуть из гнезда яйца других самок. Она обычно насиживает все собственные яйца и около половины яиц других самок. Насиживающая самка позволяет другим самкам откладывать яйца в ее гнездо, не пытаясь отогнать их. Очевидно, другие самки не связаны с ней тесным родством, так что ее терпимость нельзя объяснить отбором родичей. Самке, по-видимому, выгодно, чтобы ее кладка была окружена чужими яйцами. Хищники, совершающие набеги на гнездо (обычно шакалы и грифы), забирают лишь несколько яиц, так как каждое из них очень велико. Вероятность того, что яйца хозяйки будут похищены, снижается из-за наличия других яиц, в частности из-за того, что ее яйца собраны в середине гнезда. То же самое справедливо и в отношении самки страуса, окруженной выводком: возможно, она вынуждена заботиться о нескольких птенцах других самок, поскольку их присутствие уменьшает вероятность того, что ее собственных птенцов съедят хищники.
Альтернативная стратегия для самки страуса - сделать так, чтобы ее яйца насиживала другая самка, даже если шансы на их выживание невелики. Если откладывать большое число яиц в разные гнезда, то, возможно, такая стратегия и окупится, но она, очевидно, не является предпочитаемой стратегией, так как самки-паразиты часто остаются без самца или их потомство поедают хищники.
По причинам, обсуждавшимся в гл. 9, самцы обычно конкурируют за возможность спариться с самками. Такая конкуренция может принимать разные формы, причем большей частью это, очевидно, агрессивное соперничество. Если самец может препятствовать доступу других самцов к самкам, то ему, должно быть, нетрудно заставить самку спариваться с ним, так как у нее не остается другого выбора. У полигинных видов, таких, как благородный олень и морской слон, доминантные самцы могут собрать гарем, который они охраняют от других самцов. Агрессивное соперничество - дело рискованное, сопровождающееся ранениями или даже гибелью, поэтому в царстве животных существует много других форм соперничества самцов.
В случаях, когда самец получает доступ к самкам, прямо защищая их от соперников, проявляется тенденция к социальной организации вида, основанной на гаремной структуре. Создание гарема значительно облегчается, если самки стремятся собраться в группы, что служит защитой от хищников или облегчает использование пищевых ресурсов. У морских слонов образованию гаремов способствует то, что на берегу имеется немного мест, где животные могут спокойно отдыхать. Антилопы и олени образуют стада, чтобы защищаться от хищников. Самцы - владельцы гаремов - часто держатся с краю, защищая самок от соперников, и даже небольшое стремление самок рассеяться чрезвычайно осложнило бы жизнь стада.
У некоторых видов самцы конкурируют
не за доступ к самкам, а за оплодотворение. Иногда это называют конкуренцией спермиев. Например, самцы навозной мухи (Scatophaga stercoraria) конкурируют за самок, и иногда одному из них удается прогнать другого во время копуляции и занять его место. Паркер (Parker, 1978) показал, что, когда два самца спариваются с одной самкой, сперма второго самца оплодотворяет большую часть яиц. Паркер облучал самцов 60Со, что предотвращало развитие яиц в случае оплодотворения их спермиями этих самцов. Если облученный самец копулировал с самкой после того, как с ней спарился необлученный самец, то у самки развивалось только 20% яиц, а если до него - 80% яиц. Каким-то образом спермии второго самца вытесняют большинство спермиев первого самца.
Для копуляционного поведения грызунов характерны многократные интромиссии и эякуляции. Эти действия, По-видимому, облегчают продвижение спермы по половым путям самки. Период покоя, следующий за эякуляцией, очевидно, необходим для завершения продвижения спермиев и для успешной имплантации оплодотворенного яйца в стенку матки (Adler, 1969). Если в этот период с самкой удается копулировать другому самцу, то продвижение спермиев первого самца может быть нарушено и отцом потомства становится второй самец.
Неудивительно, что у видов, у которых возможна конкуренция спермиев, самцы принимают определенные меры предосторожности. Многие млекопитающие совокупляются в уединении, избегая возможного вмешательства. Самец навозной мухи после копуляции сидит на спине самки и охраняет ее, пока та не отложит яйца. Некоторые насекомые после копуляции замазывают половое отверстие самки. Считают, что такие копуляционные пробки предотвращают последующее совокупление у некоторых водяных жуков, дневных и ночных бабочек (Wilson, 1975). Эффективной копуляционной пробкой может служить продолжительное совокупление. Самец комнатной мухи остается в положении копуляции около часа, хотя большая часть спермы передается им самке в течение первых 15 мин. Некоторые ночные бабочки копулируют целый день. У мухи Johannseniella nitida самка съедает самца сразу после копуляции, оставляя только его гениталии, которые служат копуляционной пробкой (Wilson, 1975).
Если самец не в состоянии добиться самки в соперничестве с другими самцами, то он может копулировать украдкой или обманом. Например, самцы лягушки-быка (Rana catesbeiana) конкурируют за территорию в прудах, куда самки приходят откладывать икру. Самки предпочитают определенные участки прудов, обычно те, где вода теплая, а растительность не слишком густая. В таких условиях икринки быстро развиваются, образуя плотные шары, относительно защищенные от нападения пиявок (Howard, 1978). Самцы используют голос и физическую силу, сражаясь за лучшую территорию. Самцы, которые старше и крупнее, обычно побеждают, а небольшие молодые самцы могут остаться вообще без территории. Эти молодые самцы пользуются такой стратегией - они молча сидят около «поющего» самца и пытаются перехватить самок, которых он привлекает. Точно так же молодые морские слоны нередко пытаются присоединиться к гарему самца-доминанта, при этом ведут они себя как самки. Затем они украдкой совокупляются, пока хозяин гарема разбирается с соперником (Le Boeuf, 1974). Молодые самцы благородного оленя также могут пытаться украдкой спариваться с самками из гарема доминирующего самца.
Альтернативные стратегии внутри вида могут использоваться самцами на разных стадиях развития, как в примерах, обсуждавшихся здесь. У некоторых видов отдельные самцы могут использовать обе стратегии.
Например, у квакши (Hyla cinerea) самцы, которые громко призывают самок, и молчащие самцы-спутники, которые дожидаются возможности перехватить самку, не различаются по размерам. Примерно один из семи самцов молчит, однако их стратегия может меняться в разные ночи. У самцов-спутников успех размножения может быть почти такой же, как и у «поющих» самцов (Perril et al., 1978), а у
лягушки-быка, наоборот, успех самцов-спутников гораздо ниже, чем у доминирующих крикунов. Если предположить, что самцы одинаковых размеров свободны в выборе той или другой стратегии, то можно ожидать, что лучшая стратегия зависит от выбора других самцов. Так, если большинство самцов выбрали стратегию крикунов, тогда стратегия спутников, вероятно, принесет изрядную прибыль. Если же, однако, слишком многие самцы выберут стратегию спутников, то самок на этот участок будет привлекаться слишком мало; в таком случае лучше выбрать стратегию крикунов.
Эту ситуацию можно проанализировать в понятиях теории ЭСС. Чтобы это сделать, нам надо определить затраты на осуществление каждой стратегии и выигрыш от нее. Стратегия крикуна может оказаться невыгодной, так как крик привлекает хищников, а проигрыш при выборе стратегии спутника может быть связан с отсутствием собственного участка. Говард (Howard, 1979) установил, что кричащие лягушки-быки обычно привлекают каймановых черепах, что же касается квакши, то здесь, чтобы провести полный сравнительный анализ, нужны дополнительные сведения об условиях их обитания. Точно так же территориальные самцы полевого сверчка (Gryllus integer), которые призывают самок трением надкрыльев друг о друга, привлекают и незваных гостей (Cade, 1979) Это многочисленные самцы-спутники, которые совокупляются с приближающимися к участку самками, и паразитические мухи (Euphasiopteryx ochracea), чьи личинки развиваются в теле сверчка и убивают его в процессе своего развития. Кейд обнаружил, что поющие самцы в пять раз чаще подвергаются нападению этих паразитических мух, чем самцы-спутники, но зато и самок они оплодотворяют больше.
У многих видов есть самцы, которые никогда не становятся доминирующими и никогда не владеют территорией. Они постоянно могут принимать альтернативные стратегии. Так, самцы турухтана, спаривающиеся на. току, принимают альтернативные стратегии. Территориальные самцы терпимо относятся к светлоокрашенным самцам-сателлитам, вероятно, потому, что они привлекают самок к току. Каждый самец-сателлит затаивается вблизи конкретного участка и украдкой спаривается с самками, пока хозяин участка занят чем-нибудь другим. Тип самца в данном случае определен генетически, поэтому самцы не могут менять стратегию на другую на протяжении всей своей жизни.
Интересная ситуация наблюдается у ушастого окуня (Lepomis macrohirus), которого Гросс и Чарнов (Gross, Charnov, 1980) изучали на озере Опиникон (пров. Онтарио). Эти рыбы образуют колонии, и каждый самец, имеющий участок, строит гнездо, в которое самка откладывает икру. Самец оплодотворяет икру, охраняет гнездо и заботится о развивающихся мальках. Окраска тела у этих заботливых самцов светлая, с темной желто-оранжевой грудью. Однако можно выделить и два других типа самцов. «Самцы-воришки» имеют светлую окраску. Не привлекая внимания, они держатся у самого дна озера. Тотчас после того, как самка отнерестится, они могут быстро проникнуть в гнездо и оплодотворить часть икры. У «самцов-спутников» окраска тела темная, с еще более темными вертикальными полосами. Окраской и поведением они имитируют самку. Они могут неторопливо заплывать в гнездо и проявлять агрессивность к другим рыбам такого же размера. Гросс и Чарнов обнаружили, что эти три типа самцов становятся репродуктивно активными в разном возрасте. Они установили, что «самцы-воришки» развиваются в самцов, имитирующих самок, и становятся самцами-спутниками. Самцы обоих типов обманом добиваются доступа в гнездо к неоплодотворенной икре. У них не проявляются ни территориальное поведение, ни забота о потомстве. У территориальных самцов не наблюдается репродуктивной активности почти до семилетнего возраста. Они конкурируют друг с другом за места для гнезд, и на их родительском вкладе паразитируют самцы-воришки и самцы-спутники.
Таким образом, у самцов ушастого окуня жизненный цикл может протекать по-разному. Самец может рано стать поло-
возрелым и принять паразитическую, зависимую стратегию размножения, или половое созревание может затянуться, и он станет территориальным самцом и будет заботиться обо всей оплодотворенной икре в гнезде независимо от того, является он отцом потомства или нет. По мнению Гросса и Чарнова (Gross, Charnov, 1980), равновесное соотношение (или ЭСС) самцов, принявших ту или иную стратегию, таково, что приспособленность особи на протяжении ее жизни будет одинаковой при выборе любого образа жизни. На основе этого они подсчитали, что равновесная доля самцов в возрасте двух лет, которые выбирают образ жизни «воришек» и «спутников», должна быть равна доле икринок, оплодотворенных в каждом сезоне размножения всеми самцами популяции - «воришками» и «спутниками». Они обнаружили, что 21% самцов в возрасте двух лет развивается в «воришек». Доля потомков, полученных «воришками» и «спутниками», колеблется от 3 до 30% в зависимости от глубины водоема. Таким образом, они получили подтверждение своей модели ЭСС.
Дата добавления: 2016-06-18; просмотров: 2493;
Поиск по сайту
Узнать еще
- CRM - стратегия управления взаимоотношениями с клиентом
- I. Социально-экономическое и политическое положение Порты во второй половине XIX в.
- I. Япония во второй половине XIX в. Продолжение модернизации страны
- II. Организация дезинфекционных и стерилизационных мероприятий в организациях, осуществляющих медицинскую деятельность
- II. Функционально-структурная организация и программное обеспечение персонального компьютера
- XX съезд КПСС. Демократизация жизни страны во второй половине 50 - х годов. “Оттепель”.
- XXII. ОРГАНИЗАЦИЯ РАБОТ ПО ОБЕСПЫЛИВАНИЮ РУДНИЧНОГО ВОЗДУХА
- Автобусные маршруты и их организация

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
