Приспособленность в естественных условиях обитания
Наше обсуждение совокупной приспособленности подразумевает, что существуют два альтернативных способа описания естественного отбора. В популяционной генетике единицей отбора считается ген, и число репликаций гена максимизировано естественным отбором. Поскольку логика этого подхода весьма привлекательна, его иногда можно использовать, чтобы осветить некоторые аспекты поведения животных (Dawkins, 1976), но в действительности он не очень подходит для исследователей поведения отдельных животных. Эквивалентный подход - это считать каждое отдельное животное единицей отбора, а совокупную приспособленность - количественным показателем, максимизированным естественным отбором (Dawkins, 1978). Для практических целей успех размножения отдельной особи хорошо отражает ее совокупную приспособленность (Grafen, 1982).
Приспособленность, обусловленная поведением в природных местообитаниях, зависит от давления отбора, действующего в настоящий момент. Теоретически любой аспект поведения животного оказывает влияние на его приспособленность либо из-за прямых последствий этого поведения, либо просто потому, что один тип поведения мешает другому, который может быть более или менее выгоден. На практике природные условия обитания так сложны, что чрезвычайно трудно измерить изменения приспособленности, обусловленные поведением. Существуют два основных подхода к решению этой проблемы. Один из них - это попытка тщательно определить затраты и выигрыши, связанные с поведением в природной среде. Этот подход обсуждается в гл. 24. Другой подход - прямо измерить изменения в успехе размножения, которые вызваны определенными типами поведения.
В разд. 6.1 было показано, что значение для выживания определенных типов поведения, таких, как удаление яичной скорлупы из гнезда обыкновенной чайкой, можно оценить в экспериментах, проводимых в полевых условиях. Поскольку такие эк-
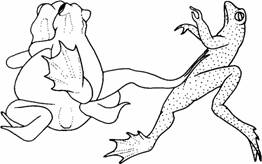
| Рис. 6.11. Спаривающиеся жабы. Самец отталкивает соперника. (Рисунок из Davies, Halliday, 1977. Фотография Tim Halliday.) | 
|
сперименты помогают понять значение конкретных морфологических и поведенческих признаков, их не связывают прямо с вопросом приспособленности. Понятие стоимости выживания обычно связано с относительно небольшим кругом вопросов, таких, как выживание молодых на протяжении конкретного сезона размножения. В идеале оценки приспособленности должны быть связаны с долговременным сохранением конкретных генетических признаков или долговременными последствиями, связанными с успехом размножения, или конкретным типом поведения.
Иногда различия в успехе размножения можно относить на счет довольно простых типов поведения. Например, Девис и Холлидей (Davis, Halliday, 1977) обнаружили, что на успех размножения пары жаб Bufo bufo влияют как размеры самки, так и степень соответствия спаривающихся особей (рис. 6.11) по размерам. Так, более крупная самка производит больше яиц, и когда спаривающиеся особи соответствуют друг другу по размерам, оплодотворяется больше яиц. Самец, который выбирает самку лишь немного крупнее, чем он сам, должен достичь наибольшего успеха в размножении, тогда как самка должна отдавать предпочтение самцу покрупнее, который мог бы отталкивать соперников. Следовательно, каждая особь (и самец, и самка) должна предпочитать партнера крупнее, чем она сама. Наблюдаемые случаи спаривания оказываются между двумя этими теоретическими крайностями, причем самки обычно крупнее самцов (рис. 6.12).
На севере шт. Мичиган изучали успех спаривания в природной популяции стре-
юзы Enallagma hageni (Fincke, 1982). Эти стрекозы размножались в мелком пруду, отделенном от озера песчаной косой шириной 10-20 м, а питались в открытой болотистой местности, примыкающей к месту размножения. В солнечные дни самцы стрекоз летели от места добывания пищи к пруду примерно с 9 ч 30 мин. К 13 часам большая часть отмеченных на пруду самцов не спаривалась. Число спаривающихся пар (соединенных вместе в виде тандема) возрастало до трех часов дня и оставалось высоким почти до половины шестого вечера. В один день почти все самки были обнаружены в тандемных парах, но обычно более половины самцов остаются без пары. Поскольку самцов в районе размножения больше, чем самок, конкуренция между самцами довольно интенсивная. Самцы перехватывают самок по дороге к пруду, и почти все добивающиеся успеха в спаривании самцы имеют пару до подлета к воде. Конкуренция между самцами происходит в форме столкновений с тандемной парой. Такое беспокойство иногда приводит к разъединению пары и замене одного самца другим.
Удивительно, но Финк (Fincke) не выявил никакой связи между размерами тела самца и его успехом в спаривании. Очевидно, маленький самец так же способен к полету в тандеме с самкой, сак и крупный. Хотя крупные самцы, быть может, более успешно вытесняют соперника из пар, чем маленькие, - это преимущество, вероятно, сводится на нет тем фактом, что приставая к тандемным парам, одиночный самец подвергается риску быть съеденным вблизи воды. Финк обнаружил, что более крупные самцы стремятся проводить большую часть времени вблизи воды, и это заставляет предполагать, что беспокойство пар и есть их основная стратегия. Однако альтернативная стратегия - поиски свободных самок, вероятно, оказывается более успешной.
Процент успешно размножавшихся самцов значительно менялся изо дня в день, причем в среднем спаривалось 39% самцов. Успех размножения на протяжении жизни также был весьма различен.
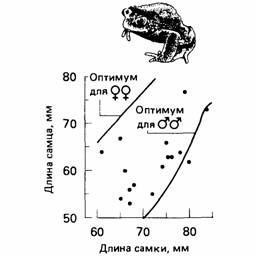
| Рис. 6.12. Оптимальное и наблюдавшееся спаривания у жаб (Bufo bufo) разных размеров. Оптимумы рассчитаны при допущении, что в сезон откладывается максимальное число оплодотворенных яиц. Кривая оптимума для самок представляет минимум, так как в эксперименте невозможно было добиться, чтобы самцы спаривались с меньшими по размерам самками. Наблюдавшиеся спаривания представлены отдельными точками. (По Davies, llallidav, 1977.) |
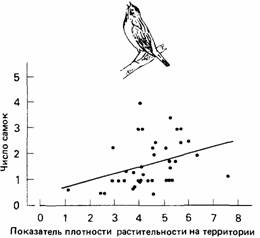
| Рис. 6.13. Число самок, привлеченных самцом овсянки американской (Spiza americana), как функция показателя плотности растительности на занимаемой им территории. (По Zimmerman, 1971.) |
Лишь 59% самцов в популяции спаривались по крайней мере однажды, и эта величина тесно связана с продолжительностью жизни. Главной причиной гибели на месте размножения было поедание стрекоз пауками, разнокрылыми стрекозами или лягушками. Таким образом, оказывается, что наиболее важный компонент успешной стратегии размножения у этого вида - избегать хищников, чтобы выжить и спариться, как только представится возможность.
Многие животные во время сезона размножения защищают территорию. В некоторых случаях - это форма распределения животных по территории, что дает возможность найти партнера для размножения (Davies, 1978). Иногда территории различаются по качеству, и можно ожидать, что самцы, которые защищают лучшие территории, добьются большего успеха в размножении. Например, у трехиглой колюшки самцы, занимающие большую территорию, привлекают больше самок (Assem, 1967). И самки, которые откладывают яйца на большой территории, размножаются успешнее благодаря тому, что меньше яиц поедают другие самцы. Самцы-соперники с соседних территорий пытаются разрушить гнездо, но и это труднее сделать, когда территория велика (Black, 1971; Wootton, 1976). Качество территории может зависеть не только от ее размеров, но и от наличия пищи и удобных мест для гнезда (рис. 6.13). В тех случаях, когда животные конкурируют из-за партнера для размножения, территории или местообитания (Partidge, 1978), преуспевающие могут быть приспособленнее в различных отношениях. Они могут добиваться большего успеха в размножении не только в результате того, что они успешнее привлекают партнеров или занимают территорию, но также и потому, что они приспособленнее в других отношениях. Это вечная проблема при оценке приспособленности. В природе так много взаимодействующих факторов, что поистине невозможно учесть их все.
ДЛЯ ЗАПОМИНАНИЯ
1. Значение для выживания конкретных типов поведения можно изучить в экспериментах, проводимых в природных условиях.
2. Изучение причин смертности в популяции зачастую может дать очевидные доказательства давления отбора.
3. Приспособленность в дарвиновском смысле измеряется числом потомков, подсчитанных на определенной стадии жизненного цикла.
4. Совокупная приспособленность особи зависит от выживания ее потомков и близких родственников. Даже если животное не оставляет потомства, его совокупная приспособленность может быть не равна нулю, поскольку его гены будут передаваться следующим поколениям племянницами, племянниками, двоюродными братьями и сестрами.
5. При изучении приспособленности в природных условиях обычно приходится пользоваться показателем успеха спаривания или успеха размножения. Эти показатели дают лишь приближенное значение приспособленности, поскольку в них не учитывается жизнеспособность потомков.
Рекомендуемая литература
Dawkins R. (1982). The Extended Phenotype, Freeman, New York. Krebs J.R., Davies N. B. (1981) An Introduction to Behavioural Ecology, Blackwell Scientific Publications, Oxford.
Дата добавления: 2016-06-18; просмотров: 1985;
Поиск по сайту
Узнать еще
- Cдвиг ветра. Воздействие на взлёт и посадку ВС. Рекомендации по выполнению полётов и управлению воздушным движением в условиях сдвига ветра.
- Абсолютная мобильность капитала в условиях фиксированных валютных курсов
- АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ
- АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ И ЕЕ ЗНАЧЕНИЯ ДЛЯ МЕДИЦИНЫ
- Аналитическая оценка характеристик антенного окна в условиях нагрева
- Анаэробный распад загрязнений в искусственных условиях
- Аэробный распад загрязнений в естественных условиях
- Аэробный распад загрязнений в искусственных условиях

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
