Рецептор; 2 – афферентный нейрон; 3 – вставочный нейрон; 4 – мотонейрон; 5 – эффектор (скелетная мышца).
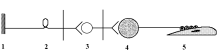 (СНС)
(СНС)
1 – рецептор; 2 – афферентный нейрон; 3 – центральный (преганглионарный) нейрон; 4 – ганглионарный нейрон (симпатический ганглий); 5 – эффектор (гладкая мышца).
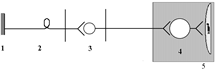 (ПСНС)
(ПСНС)
1 – рецептор; 2 – афферентный нейрон; 3 – центральный (преганглионарный) нейрон; 4 – ганглионарный нейрон (парасимпатический ганглий); 5 – эффектор (гладкая мышца).
12. Понятие о рефлексе. Строение рефлекторной дуги и функции его звеньев. Нарисовать схему рефлекторной дуги вегетативного рефлекса.(см. выше)
Рефлекс – ответная реакция организма на раздражение рецепторов, осуществляемая при обязательном участии нервной системы
Первое звено (рецептор) воспринимает раздражение, трансформируя энергию раздражения в нервный импульс. Второе звено (афферентный нейрон) проводит импульсы в ЦНС. Третье звено Вставочные нейроны – передают импульсы к эфферентному нейрону и обеспечивают связь данной рефлекторной дуги с другими отделами ЦНС. Четвертое звено (эфферентный нейрон) перерабатывает информацию, поступающую к нему от вставочных нейронов ЦНС и формирует ответ в виде нервных импульсов, посылаемых к 5-у звену – к рабочему органу.
13. Проприорецепторы, их виды, функции, механизмы возбуждения.
Проприорецепторы-рецепторы опорнодвигательного аппарата(мышц, сухожилий, фасций, суставов), что принимают участие во всех актах, что связаны с работой мышц, суставов и сухожилий.
 К проприорецепторам относят мешочные веретена и сухожильные рецепторы (Гольджи). Функции: принимают участие в осознание направления и скорости движения конечностей, определить положение отдельных частей тела в пространстве, построение трехмерного чувствительного окружающего мира(стереогноз).
К проприорецепторам относят мешочные веретена и сухожильные рецепторы (Гольджи). Функции: принимают участие в осознание направления и скорости движения конечностей, определить положение отдельных частей тела в пространстве, построение трехмерного чувствительного окружающего мира(стереогноз).
Мышечные веретена- расположены параллельно к мышечным волокнам, и при расслаблении мышцы, веретена растягиваются. Тем самым учащая ПД в афферентном нейторе возбуждают свой центр, тормозя нейроны центра антагониста
Рецепторы Гольджи- расположены между сухожильными волокнами мышц(последовательно), при напряжение мышцы, сухожилья растягиваются, деформируя рецептор, в следствии чего возрастает частота ПД в афферентном нейроне, что образует синапс с тормозными интернейронами, тем самым тормозит гомономные мотонейроты.
14.Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных синапсах.
Механізм передачі збудження через центральний аксосоматичний хімічний синапс полягає в наступному: ПД поширюється по мембрані аксона далі по мембрані пресинаптичній підвищення проникності пресинаптичної мембрани для іонів Сa2+ вхід їх в нервове закінчення за градієнтом концентрації вихід медіатора(АХ/НА) в синаптичну щілину дифузія медіатора до постсинаптичної мембрани взаємодія з мембранними циторецепторами(N-холино/адрено) збільшення проникності постсинаптичної мембрани для іонів Na+ вхід іонів Na+ в тіло клітини через постсинаптичну мембрану деполяризація мембрани (ЗПСП місцеве збудження) ЗПСП як місцеве збудження поширюється на сусудні ділянки постсинаптичної мембрани та мембрани аксонного горбика з допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тому викликають деполяризацію мембрани. Цікавим є виникнення місцевих струмів між постсинаптичною мембраною (там ЗПСП) та мембраною аксонного горбика – початковий сегмент аксона, мембрана якого має найбільшу збудливість, поріг деполяризації (ΔЕ) там складає 10-15 мВ. Тому ПД виникає під впливом місцевих струмів саме там. Це відбувається, якщо під впливом місцевих струмів деполяризація мембрани аксонного горбика досягає критичного рівня виникнення серії ПД ритмічний розряд нейрона.
Особливості передачі збудження через центральні аксо-соматичні хімічні синапси.
1. Одностороннє проведення.
2. Сповільнення проведення – характеризується наявністю синаптичної затримки – час від виникнення ПД на пресинаптичній мембрані до виникнення ПД на мембрані аксонного горбика. Воно складає 2-3 мс.
3. Швидке порушення проведення через виснаження запасів медіатора.
Ці три особливості характерні для будь-яких хімічних синапсів, бо синаптична затримка та швидкість розвитку втоми в центральних синапсах більша, ніж в нервово-м’язових. Наступні дві особливості характерні лише для центральних синапсів.
4. Один ПД через центральний синапс не проходить тому, що зумовлює на постсинаптичній мембрані виникнення одного ЗПСП, який має амплітуду 1 мВ та тривалість 15 мс. Оскільки поріг деполяризації аксонного горбика складає 10-15 мВ, один ЗПСП, що поширився на мембрану аксонного горбика, не може викликати деполяризацію цієї мембрани до критичного рівня.
5. Умовою передачі збудження через центральний нервовий синапс є сумація ЗПСП на тілі нейрона.
До медіаторів, що викликають деполяризацію постсинаптичної мембрани та приймають участь в передачі збудження, відносять: ацетилхолін, норадреналін, серотонін та багато інших.
15.Торможение в ЦНС. Его виды и физиологическая роль. Механизмы и закономерности пре- и постсинаптического торможения. Нарисовать схемы пре- и постсинаптического торможения.
Торможение- нервный процесс, что обусловлен возбуждением и внешне проявляется угнетением другого возбуждения
| За локализацией | За характером поляризации | За строением тормозных нейронных цепей |
| -пресинаптическое -постсинаптическое | -гиперполяризационное -деполяризационное | -реципрокное -возвратное -латеральное |
Торможение является важным фактором координационной деятельности ЦНС, участвует в обработке информации, поступающей к нейрону, выполняет охранительную роль.
Пресинаптическое торможение-связано с угнетением проведения нервных импульсов аксональных(пресинаптических) окончаниях. К возбужденному аксону подходит вставочный тормозной аксон что выделяет тормозной медиатор ГАМК
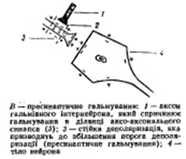 Постсинаптическое торможение- обусловлено выделением из пресинаптического окончания аксона тормозного медиатора, что понижает или тормозит возбудимостьсомы иди дендридов(аксо-сомальный, аско-дендритный синапс)
Постсинаптическое торможение- обусловлено выделением из пресинаптического окончания аксона тормозного медиатора, что понижает или тормозит возбудимостьсомы иди дендридов(аксо-сомальный, аско-дендритный синапс)

16. Суммация возбуждения и торможения нейронами ЦНС
Суммация возбуждающих постсинаптических потенциалов (ВПСП) или тормозных постсинаптических потенциалов (ТПСП) в нейронах нервного центра, в результате которых может возникнуть импульсное возбуждение.
Различают:
а)временную- при поступлении серии нервных импульсов по одному и тому же афферентному пути.
Б)пространственную-при которой импульсы подходят одновременно по многим афферентным волокнам.
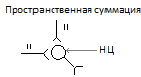
17.Проводниковая функция СМ. Спинальный шок.

 Спинальный шок - резкое угнетение возбудимости и рефлекторной деятельности спинного мозга ниже места его травмы или перерезки. Возникает вследствие выключения активирующего влияния вышележащих отделов ЦНС на спинной мозг.
Спинальный шок - резкое угнетение возбудимости и рефлекторной деятельности спинного мозга ниже места его травмы или перерезки. Возникает вследствие выключения активирующего влияния вышележащих отделов ЦНС на спинной мозг.
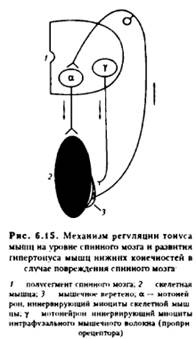
18. Фазные и тонические рефлексы СМ, их рефлекторные дуги и физиологическое значение.
Фазные р-сы – однократное сгибание(разгибание) конечности при однократном раздражении кожи или мышечных рецепторов. (участвуют в акте ходьбы)
Тонические р-сы – р. что обеспечивают сохранение положения тела в пространстве, взаимное расположениеего частей, равновесие тела, конечностей и глаз с положением головы, включает статические и статокинетические рефлексы.
19.Двигательные рефлексы заднего и среднего мозга, децекебральная ригидность.
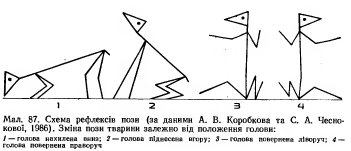
Сгибательные, разгибательные, ритмические, познотонические
Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу
 20.РФ ствола мозга, механизм поддержания ее тонуса. Нисходящие и восходящие влияния РФ.
20.РФ ствола мозга, механизм поддержания ее тонуса. Нисходящие и восходящие влияния РФ.
РФ - скопление нейронов различных типов и размеров, связанных множеством волокон, идущих в различных направлениях и образующих сеть на всем протяжении ствола мозга, а также в шейных и верхнегрудных сегментах спинного мозга. Для поддержания тонуса РФ получает импульсы от всех рецепторов организма и от всех отделов ЦНС. Нейроны РФ являются полимодальными, посылают импульсы ко всем отделам ЦНС.
Аферентних шляхів РФ дуже багато, вона отримує інформацію від:
- усіх специфічних висхідних еферентних шляхів, які віддають колатералі до ядер РФ, заходячи в стовбур мозка;
- від іних утворень ЦНС, а саме:
-від кори головного мозку;
-від гіпоталамуса;
-від мозочка;
-від базальних ядер;
-від інших утворень.
Еферентні зв’язки РФ можна поділити на висхідні та низхідні.
Висхідні зв’язки РФ та висхідні впливи:
- через неспецифічні ядра таламуса передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини. Цей вплив забезпечує бодьорий стан організму та визначає рівень активності клітин кори. РФ має окремі структури (гіпногенні зони), котрі зменшують ступінь (рівень) активності клітин кори та зумовлюють стан сну;
- по ретикулогіпоталамічним шляхам передає інформацію від різних рецепторів в гіпоталамус.
Характер низхідних ретикулоспінальних впливів:
- від медіального ядра РФ заднього мозку починається ретикулоспінальний (входить до складу МНС) – специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок;
- від інших ядер РФ беруть початок шляхи, які неспецифічно активують чи гальмують спінальні мотонейрони і таким чином визначають ступінь їхньої активності;
- від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспінальним шляхам передається інформація до симпатичних рухових нейронів спинного мозку при регуляції відповідних функцій організму.
21. Роль среднего мозга в обеспечении положения тела. Двигательные рефлексы среднего мозга.
Середній мозок (СрМ) за участі сітчастої речовини опрацьовує аферентну інформацію, яка поступає в спинний та задній мозок. Нова інформація поступає в СрМ від зорових та слухових рецепторів. Її опрацювання відбувається в ядрах передніх (від зорових рецепторів) та нижніх (від слухових рецепторів) горбків. На основі опрацьовання інформації від усіх цих рецепторів СрМ здійснює контроль за станом зовнішнього та внутрішнього середовища організма.
Важливими надсегментарними руховими ядрами СрМ є:
1) червоні ядра – від них інформація від нейронів спинного мозку передається по шляхах що перехрещуються (руброспінальні шляхи – елемент ЛНС);
2) ретикулярна формація;
3) чорна субстанція – знаходиться в середньому мозку, але функціонує як єдине ціле з базальними ядрами.
Руброспінальні впливи характеризуються активацією - та -мотонейронів флексорів та гальмуванням - та -мотонейронів екстензорів. Ці впливи протилежні вестибулоспінальним. Завдяки руброспінальним впливам тонус м’язів мезенцефальної тварини більш рівномірніший, ніж у децереброваної (тонус флексорів та екстензорів більш збалансований). Тому, поза мезенцефальної тварини нормальна. Тобто, посилена антигравітаційна поза децереброваної тварини перетворюється в нормальну у мезенцефальної завдяки руброспінальним впливам.
За участю рухових ядер СрМ здійснюються позо-тонічні рефлекси двох видів:
1.Випрямні – забезпечують відновлення порушеної пози. У тварин, що мають шию, випрямні рефлекси мають 2 фази:
а) Рефлекс випрямлення голови – він починається з подразнення отолітових вестибулорецепторів у відповідь на порушення нормального положення голови в просторі (наприклад при падінні). Рівень замикання цього рефлексу – СрМ. Для його здійснення необхідна участь червоних ядер та РФ СрМ. По низхідних шляхах від цих структур інформація передається до мотонейронів спинного мозку перерозподіл тонусу м’язів шиї голова набуває нормальне положення в просторі (тім’ячком доверху).
б) Рефлекс випрямлення тулуба – починається з подразнення пропріорецепторів шиї, що спричиняється рефлексом випрямлення голови, так як при цьому відновлюється нормальне положення голови у просторі, але порушується її нормальне положення відносно тулуба. Вхід інформації від пропріорецепторів шиї здійснюється на рівні спинного мозку підйом до надсегментарних структур СрМ обробка цієї інформації передача до мотонейронів спинного мозку до м’язів, які забезпечують відновлення нормального положення тулуба відносно голови у просторі.
2. Стато-кінетичні – забезпечують збереження пози рівноваги при русі з прискоренням. При русі з лінійним прискоренням збуджуються отолітові рецептори, а при русі з кутовим прискоренням – рецептори півколових канальців. Вхід цієї інформації в ЦНС здійснюється на рівні заднього мозку підйом до структур СрМ обробка та передача низхідними шляхами до мотонейронів спинного мозку перерозподіл тонусу м’язів, який забезпечує збереження рівноваги під час руху з прискоренням. Важливим компонентом стато-кінетичних рефлексів є присінково-окорухові рефлекси ністагм очей більш продовжена фіксація предметів в полі зору під час руху краща орієнтація в просторі більш надійне збереження пози при русі.
Ще раз підкреслимо, що рівень замикання випрямних та стато-кінетичних рефлексів – СрМ. Тому, їх можливо отримати у мезенцефальної тварини і неможливо у децереброваної чи спінальної.
За рахунок обробки інформації від зорових та слухових рецепторів (за участю ядер чотирьох горбків) рухові ядра СрМ забезпечують здійснення орієнтаційних рефлексів – у відповідь на сильні звукові та світлові подразники мезенцефальна тварина повертає голову в бік подразника, насторожує вуха, в неї дещо посилюється тонус флексорів – тварина ніби готується до відповіді на цей подразник.
Роль СрМ в здійсненні фазичних реакцій залежить від рівня організації тварини:
- мезенцефальні земноводні (жаба) здатні до локомоцій – руху у просторі (стрибають, плавають);
- кішки та собаки не здатні до локомоцій, але здійснюють локомоторні синергії – злагоджені рухи кінцівок, що нагадують акт ходьби: їх інтегрують структури СрМ разом з структурами заднього та спинного мозку.
У людини СрМ не забезпечує формування нормальної антигравітаційної пози в спокої та підтримання її при русі. Для здійснення всіх цих функцій у людини необхідна участь кори великих півкуль (принцип кортикалізації функцій).
22.Мозжечок, его функции. Симптомы поражения.
Мозочок (Мз) відіграє дуже важливу роль в координації рухів і виконує цю функцію на основі своїх аферентних та еферентних зв’язків.
По аферентним волокнам сигнали поступають в кору мозочка, яка має складну будову та забезпечує тонку обробку поступившої інформації. Причому, кінцеву обробку інформації здійснюють грушоподібні клітини Пуркіньє – по відростках цих клітин (по їх аксонах) здійснюється еферентний вихід обробленої інформації з кори Мз. Далі, інформація здебільшого прямує до ядер Мз по аксонах їх нейронів здійснюється вихід інформації за межі Мз.
Аферентні зв’язки Мз:
1.Від рецепторів шкіри та пропріорецепторів Мз отримує інформацію по спіно-церебелярних шляхах; ця інформація забезпечує можливість контролю за рухом який здійснюється.
2.Від вестибулорецепторів (через вестибулярні ядра) – контроль за збереженням рівноваги при русі.
3.Від всіх рухових ядер стовбуру (ретикулярна формація, краєві ядра).
4.Від базальних ядер.
5.Від моторної і асоціативної кори (від моторної кори надходить копія команди до виконання руху, яка відправляється до мотонейронів; від асоціативної кори – інформація про ціль руху).
6.Від зорових та слухових рецепторів – контроль за рухами, які відбуваються.
Еферентні зв’язки Мз:
1.З руховими ядрами стовбуру (ретикулярна формація, вестибулярні ядра, червоні ядра), через які Мз здійснює вплив на мотонейрони і на м’язи.
2.З базальними ядрами.
3.З асоціативною та моторною корою
4.З гіпоталамусом.
Мз отримує інформацію про характер руху від рецепторів і при необхідності може швидко корегувати ці рухи та їх програму. При цьому різні відділи Мз здійснюють корекцію різних рухів.
Древній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра до медіального ретикулярного ядра до мотонейронів, що іннервують м’язи тулуба та кінцівок.
Таким чином, кора древнього та старого Мз, отримуючи інформацію від пропріорецепторів, рецепторів шкіри про характер руху; від вестибулорецепторів – про збереження рівноваги, і при необхідності вносить корективи в грубі позо-тонічні рухи, чому передує передача інформації по шляхах МНС, які регулюють стан м’язів тулуба і проксимальних відділів кінцівок. Ця корекція дуже важлива для підтримання рівноваги (збереження пози) при русі.
Проміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи:
а)в рух, який здійснюється (виходячи на мотонейрони спинного мозку через стовбурові рухові ядра);
б)в рухову програму, передаючи інформацію по церебело-таламо-кортикальному шляху в рухову кору.
Вихід до рухової кори і до стовбурових рухових ядер із кори проміжної частини Мз здійснюється через вставне ядро.
За рахунок корекції рухів та програми Мз добивається їх повного співпадання (їх відповідності один одному).
Кора півкуль Мз пов’язана з базальними гангліями і з асоціативною корою. Завдяки цим зв’язкам вони формують програми рухів і Мз через зубчасте ядро передає їх в рухову кору для виконання.
Основні симптоми ураження Мз у людини. Мехпнізми їх виникнення.
1.Дистонія – порушення тонусу м’язів, що спричиняється порушенням впливу Мз на стовбурові рухові ядра (ретикулярна формація, вестибулярні ядра) на мотонейрони спинного мозку на м’язи тулуба та проксимальних відділів кінцівок.
2.Дизеквілібрія – перушення рівноваги. Пов’язана з випаданням коригуючих впливів Мз на вестибулярні ядра Дейтерса.
3.Атаксія – невпорядкованість рухів. Проявляється у вигляді:
а) дисметрія – порушення амплітуди рухів;
б) порушення направленості рухів.
В основі атаксії лежать порушення координації м’язів агоністів та антагоністів.
4.Асинергія – порушення злагодженості рухів. В основі її лежить порушення координації скорочення м’язів-агоністів.
5.Астенія – швидка втомлюваність. Вона пов’язана з виконанням великої кількості “зайвих” рухів для досягнення мети.
6.Адіадохокінез – порушення синхронного виконання рухів, іх послідовності та швидкості.
7.Дисметрія – порушення рівномірності і амплітуди рухів.
8.Дисартрія – порушення артикуляції мови.
23.Таламус, его функции
Таламус є структурою проміжного мозку і виконує, головним чином, сенсорні функції – за виключенням нюхового, в тамусі переключаються всі сенсорні шляхи, які йдуть в КГМ.
В таламусі є біля 40 ядер, які розрізняються по своїм функціям. Серед них виділяють:
1. Сенсорні перемикаючі (специфічні) ядра – вони отримують інформацію від специфічних сенсорних шляхів, переробляють її і передають в сенсорні зони КГМ. Разом з цими зонами специфічні сенсорні перемикаючі ядра таламуса беруть участь в формуванні відчуттів.
2.Неспецифічні – вони отримують інформацію від ретикулярної формації стовбура мозку по шляхах больової чутливості. Вони передають інформацію до всіх зон КГМ, здійснюючи на неї неспецифічний активуючий вплив.
3.Асоціативні – отримують інформацію від специфічних сенсорних перемикаючих ядер і від неспецифічних ядер таламуса. Вони передають інформацію в асоціативні зони КГМ (фронтальна, тім’яна, вискова). Разом з даними ділянками кори, асоціативні ядра таламуса приймають участь в розпізнаванні образів.
4.Неспецифічні перемикаючі:
а)лімбічні – отримують інформацію від гіпоталамуса та передають її в лімбічну кору. Вони беруть участь у формуванні емоцій та мотивацій.
б)рухові – отримують інформацію від базальних гангліїв, мозочка, стовбурових рухових ядер і передають її в рухову кору. Беруть участь в забезпеченні рухових реакцій організму.
24. Базальные ядра ,их функции, симптомы поражения.
Базальні ядра знаходяться в глибині кінцевого мозку. До них відносяться:
- хвостате ядро;
- шкаралупа (разом з попереднім утворює смугасте тіло);
- бліда куля.
Як єдине ціле з базальними ядрами функціонують чорна субстанція та субталамічне ядро.
Ці ядра об’єднані між собою двосторонніми зв’язками, отримують інформацію від кори (асоціативних та рухових зон) та мозочка. Після відповідної обробки інформація від базальних ядер передається:
- через моторні (передні) ядра таламуса до рухової кори;
- через стовбурові рухові ядра (червоне ядро, вестибулярне ядро, ретикулярна формація) до мотонейронів спинного мозку до м’язів.
 В діяльності базальних ядер важливу роль відіграє рух збудження, який має назву циклу шкаралупи та циклу хвостатого ядра.
В діяльності базальних ядер важливу роль відіграє рух збудження, який має назву циклу шкаралупи та циклу хвостатого ядра.
1. Цикл шкаралупи – забезпечує участь базальних ядер в реалізації програм складних набутих рухових реакцій. Цикл починається з передачі інформації від премоторної зони кори (ПМК) до шкаралупи далі до блідої кулі (БК) через чорну субстанцію (ЧС) та субталамічне ядро (СТЯ) інформація, перероблена в базальних ядрах передається в першу моторну зону кори (МК) через моторні ядра таламуса.
Цикл хвостатого ядра – забезпечує участь базальних ядер в формуванні програм складних рухових реакцій. При здійсненні цього циклу інформація від асоціативних зон кори (АЗК) про те, яким повинен бути результат рухової реакції, направляється в хвостате ядро (ХЯ) далі в шкаралупу (Ш) і бліду кулю (БК) і далі через моторні ядра таламуса в рухову кору. Завдяки цьому рухова кора отримує готову рухову програму, яка підлягає наступній реалізації.
Необхідно пам’ятати, що у створенні програм рухових актів окрім базальних ядер важливу роль відіграє кора мозочка та його зубчасті ядра.
Симптоми ураження базальних ядер:
1.При ураженні смугастого тіла:
-гіпотонія м’язів;
-гіперкінези, в тому числі у спокої;
2.При ураженні блідої кулі та чорної субстанції:
-гіпокінезія, рухи втрачають індивідуальне забарвлення за рахунок зникнення міміки та пантоміміки;
-воскова ригідність – людина може довго утримувати ненормальну (незручну) позу, положення кінцівок;
-тремор спокою.
25Понятие о лимбической системе. Гипоталямус его связи и функции.
До складу лімбічної системи входять:
1.Лімбічна (стародавня і стара) кора:
-гіпокамп;
-гіпокампова закрутка (звивина);
-поясна закрутка;
-нюховий мозок (нюхові цибулини, горбки, ділянки кори над мигдалинами).
2.Підкіркові утворення:
-мигдалеподібний комплекс;
-лімбічні ядра таламуса;
-ядра перегородки;
-гіпоталамус;
3.Лімбічна частина ретикулярної формації середнього мозку.
Структури ЛС пов’язані між собою чисельними кільцевими зв’язками, що забезпечує можливість тривалої рециркуляції збудження в системі.
ЛС отримує інформацію практично від усіх рецепторів організму за рахунок аферентних зв’язків з:
-лімбічних ядер таламуса;
-ретикулярної формації середнього мозку;
-чисельних рецепторів гіпоталамуса (осмо-, термо-, глюко-, тощо).
Всі ці структури отримують інформацію від усіх рецепторів, окрім нюхових.
Такий обширний аферентний вхід дозволяє ЛС контролювати стан організму і ситуацію, в якій він знаходиться.
Лімбічна система забезпечує:
1.Формування мотивацій – стан організму, що виникає на базі потреби і змушує здійснювати реакцію, спрямовану на задоволення даної потреби.
2.Формування емоцій – стан організму, що виникає на базі потреби (з урахуванням можливості її задоволення) і мобілізують можливості організму для задоволення даної потреби, а також дозволяють швидко оцінювати інформацію, що надходить з точки зору її значення для задоволення даної потреби.
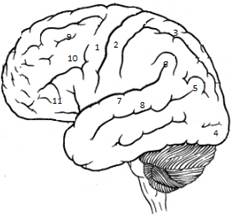
26.Сенсорные, ассоциативные и моторные зоны коры ГМ и их функции.
Сенсорні, асоціативні, моторні зони кори формують нову кору – неокортекс.
Сенсорні зони кори відповідають представництву окремих сенсорних систем (аналізаторів) у різних ділянках кори.
Так, кіркове представництво зорового аналізатора локалізується у потиличній зоні кори (шпорна закрутка), слухового – у висковій зоні, соматосенсорного – у постцентральній закрутці.
Сенсорні зони кори, діючи разом з іншими елементами специфічних каналів зв’язку в сенсорних системах, забезпечують формування відчуттів.
Асоціативні зони кори – лобна, тім’яна, вискова забезпечують виконання вищих кіркових функцій, а саме:
1.Розпізнавання образів (разом з асоціативними ядрами таламуса);
2.Здійснення абстрактного мислення, мови;
3.Беруть участь у формуванні пам’яті
4.Забезпечують вибір цілей рухових реакцій та на основі цього приймають участь у формуванні та виборі конкретних програм цих рухів.
5.Приймають участь у формуванні, зберіганні та реализації вроджених та набутих програм рухових реакцій організму.
Моторні зони – їх декілька. Головна моторна зона знаходиться у прецентральній звивині, премоторна зона – попереду головної (первинної) моторної зони і в глибині сільвієвої борозни. Моторні нейрони ї також у зоні представництва соматосенсорного аналізатора – постцентральна закрутка.
1-предцентральная/движение
2-постцентральная/ чувствительность
3-верхняя теменная/стереогноз
4-затылочная/зрение
5-угловая/чтение
6-краевая/праксия
7-верхняя височная/слуховой анализатор
8-центр Вернике(пониманае речи)
9-письмо
10-поворот глаз противоположно повороту головы
11-центр Брока(артикуляция речи)
27.Электрическая активность ГМ , частотно- амплитудная характеристика основных ритмов ЭЭГ. Клиническое значение ЭЭГ.
Электроэнцефалография - регистрация суммарной электрической активности головного мозга с поверхности кожи головы. ЭЭГ – кривая записи суммарной электрической активности головного мозга: альфа-, бета-, тэта- и дельта-волны.
Альфа-волны. Частота 8 – 13 Гц, амплитуда 25 – 50 мкВ. Закрыты глаза
Бета-волны. Частота 14 – 30 Гц, амплитуда 20 – 25 мкВ. Открыты глаза
Тэта-волны Частота 4 –7 Гц, амплитуда 100 –300 мкВ. Переход к сну
Дельта-волны Частота 0.5 – 3.5 Гц, амплитуда 100 –300 мкВ глубокий сон
28.Структурно-функциональная организация СНС : НЦ, медиаторы, специальные рецепторы, вторичные мессенджеры.
НЦ- nuc. Intermedius lateralis C8 – L2-3
Преганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы – Н-холинорецепторы
Постганглионарные: медиаторы – А, НА , ГАМК, серотонин(моноаминооксидаза)
Рецепторы:
- α1-гладкие мышцы(сужение сосудов, сокращение миометрия)
- α2- (снижает выделение медиаторов)
- β1-сердце (^ ЧCC)
- β2-гладкая мускулатура(расслабляет коронарные артерии, а. бронхов, ГМ, мочевого) Курареподобные в-ва/ бензогексоний
29.Влияние СНС на функции организма
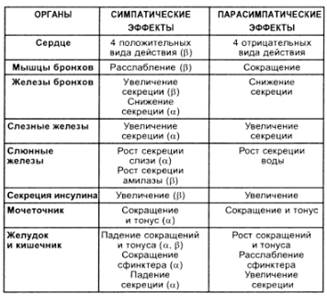
Активація симпатичної нервової системи в організмі відбувається у стані напруження, коли необхідно забезпечити мобілізацію функцій для вирішення важливої задачі (фізична робота, емоційний стрес). Симпатична нервова система забезпечує ерготропну перебудову функцій організму, а саме:
- збільшує частоту та силу серцевих скорочень;
- звужує більшість судин (такі впливи на серце і судини призводить до збільшення системного артеріального тиску – САД);
- розширює бронхи створюються умови для оптимальної вентиляції легень;
- пригнічує секреторну та рухову активність органів травної системи (функції цієї системи в стані напруження є вторинними);
- збільшує дієздатність скелетних м’язів (через зміну обміну речовин та кровотоку в м’язах);
- стимулює виділення гормонів мозкової речовини наднирників;
- збільшує збудливість рецепторів і центрів, дієздатність клітин кори головного мозку;
- змінює обмін речовин в організмі (стимулює глікогеноліз і ліполіз розпад глікогену та жирів збільшення концентрації в крові глюкози та жирних кислот субстратне забезпечення посиленого функціонування скелетних м’язів і клітин ЦНС).
-расширяет зрачок.
30. Строение ПСНС : синапсы, медиаторы, циторецепторы, блокаторы.
НЦ- nuc. Intermedius medialis S2-3(на границе передних и задних рогов)
Кранио-бульбарный(3,7,9,10)
Преганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы – Н-холинорецепторы
Постганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы: -М- холинорецепторы
Курареподобные в-ва / бензогексоний/ атропин
31. Влияние ПСНС на функции организма.
Парасимпатична нервова система активується після стану напруження і забезпечує трофотропну перебудову функцій організму, завдяки якій проходить відновлення пластичних та енергетичних ресурсів організму, що витратились під час стану напруження.
Вплив парасимпатичної нервової системи:
- зменшення ЧСС перехід серця на режим економної діяльності активний перебіг відновлювальних процесів в міокарді;
- звуження бронхів;
- активація секреторної і рухової активності органів травної системи сприятливі умови для перетравлювання та всмоктування поживних речовин посилення відновлювальних процесів;
- посилення секреції інсуліна посилення утилізації глюкози клітинами посилення відновлювальних процесів.
32.Роль центров СМ, продолговатого, среднего мозга в регуляции вегететивной функции.
Спинний мозок. Первинні симпатичні та парасимпатичні центри бокових рогів спинного мозку забезпечують реалізацію елементарних рефлексів. Парасимпатичні рефлекси (дефекація, сечовипускання, статеві) у людини в значному ступені контролюються корою головного мозку. Реалізація симпатичних спінальних рефлексів (судинно-рухових, потовидільних) перебуває під контролем сітчастої речовини заднього мозку.
Задній мозок. Вміщує в собі первинні парасимпатичні центри у вигляді відповідних ядер VII, ІX (регуляція слиновиділення) та X пар ЧМН (регуляція рухової та секреторної функції органів травної системи, роботу серця, тонус бронхіальної мускулатури). Окрім того, структури сітчастої речовини заднього мозку утворюють суднно-руховий, кардіостимулюючий та дихальний центри. Саме тому, роль заднього мозку в регуляції вегетативних функцій дуже велика. Цей рівень є першим, що може забезпечити зміну вегетативних функцій в інтересах цілісного організму (наприклад, при фізичній роботі) тут вже розміщені інтегративні центри, як приклад – сітчаста речовина.
Середній мозок. Вміщує в собі вегетативне ядро ІІІ пари чмн (ядро Якубовича), за участю якого здійснюються зіничний рефлекс (звуження зіниці) та рефлекс акомодації (зміна кривизни кришталика). Тобто, цей центр приймає участь в забезпеченні умов для ясного (чіткого) бачення.
33. Роль гипоталамуса , лимбической системы, неокортекса в регуляции вегететивных функций.
Гіпоталамус – структура проміжного мозку. Він виконує важливі функції завдяки наявності в ньому таких особливих структур:
1. В його склад входять інтегративні центри, що контролюють стан (та інтегрують):
- симпатичних центрів, а саме, сітчаста речовина (це здійснюють переважно ядра заднього гіпоталамуса);
- первинних парасимпатичних центрів стовбура мозку та спинного мозку. Саме тому гіпоталамус інтегрує нервові механізми регуляції вегетативних функцій (симпатичні та парасимпатичні) між собою.
2. До складу медіального гіпоталамуса входять нейросекреторні клітини, що обробляють інформацію як будь-які інші нервові клітини (шляхом сумації збуджень та гальмувань), та виділяють із нервових закінчень не медіатори (час життя, яких короткий, бо вони швидко руйнуються відповідними ферментами), а більш стійкі гормони:
а) Ліберини та статини, які по системі ворітних судин поступають в аденогіпофіз (передня та середня долі) і стимулюють (ліберини) чи гальмують (статини) виділення відповідних гормонів:
| Гормони гіпофіза. | Гормони гіпоталамуса. | |
| Ліберини. | Статини. | |
| Соматотропний | Соматоліберин. | Соматостатин. |
| Тиреотропний. | Тиреоліберин. | --- |
| Адренокорти. | Кортиколіберин. | --- |
| Гонадотропний. | Гонадоліберин. | --- |
| Пролактин. | Пролактоліберин. | Пролактостатин. |
| Меланоцито-ст. | Меланоліберин. | Меланостатин. |
б) Нейросекреторні клітини супраоптичного та паравентрикулярного ядер виділяють гормони вазопресин та окситоцин, які по аксонах цих нейронів надходять в нейрогіпофіз (задня доля), а звідти вже виділяються в кров.
Тропні гормони аденогіпофіза (тиреотропний, адренокортикотропний, гонадотропний) стимулюють виділення відповідних гормонів ендокринними залозами організму (щитовидною, кірковим шаром наднирників, статевими).
Тому, гіпоталамус виступає структурою, що інтегрують гуморальні (ендокринні) механізми регуляції вегетативних функцій.
Наявність в гіпоталамусі центрів, що інтегрують нервові мехінізми регуляції вегетативних функцій, а також нейросекреторних клітин, дають йому змогу інтегрувати нервові та гуморальні механізми регуляції вегетативних функцій.
Лімбічна кора (стара та древня) приймає участь в регуляції вегетативних функцій при формуванні мотивацій та емоцій.
Нова кора за участю лімбічної системи та гіпоталамуса бере участь в регуляції вегетативних функцій організму при здійсненні різноманітних реакцій поведінки.
Дата добавления: 2016-06-09; просмотров: 2234;
Поиск по сайту
Узнать еще
- N в процессе иммунного ответа антигенам 1-го класса принадлежит ведущая роль во взаимодействии между клеткой-эффектором (Т киллеры) и клеткой-«мишенью»
- Афферентный механизм произвольных движений afferentis — «приносящий»
- Афферентный сигнал. Афферентный нерв. Исполнительные органы. Обратная афферентация ( связь ).
- В-система иммунитета – это система органов, клеток и эффекторных молекул, осуществляющих гуморальную форму иммунного реагирования.
- Нервно-мышечный синапс (мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне.
- Характеристика эффекторных форм Т-лимфоцитов
- Эффекторная функция антител различных классов

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
