Немембранные органоиды
Что такое Органоиды клетки
Органоиды клетки
Органоиды, или органеллы, - постоянные специфические структуры цитоплазмы, выполняющие определенные функции, необходимые для поддержания жизнедеятельности клетки.
Различают органоиды общего значения и специальные органоиды. Органоиды общего значения имеются во всех клетках и выполняют функции, общие для всех клеток – митохондрии, рибосомы, эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и клеточный центр.
Органоиды специального значения имеются только в клетках какого-то определенного типа и обеспечивают выполнение функций, присущих только этим клеткам.
Мембранные органоиды:
- ядро;
- эндоплазматическая сеть;
- аппарат Гольджи;
- митохондии;
- лизосомы;
- пероксисомы;
- вакуоли.
Немембранные органоиды
- рибосомы;
- клеточный центр;
- цитоскелет.
Эндоплазматическая сеть открыта Портером в 1945 году. Морфология – сеть канальцев и цистерн сложенных мембранами. Различают гранулярную (шероховатую, зернистую) и гладкую ЭПС.
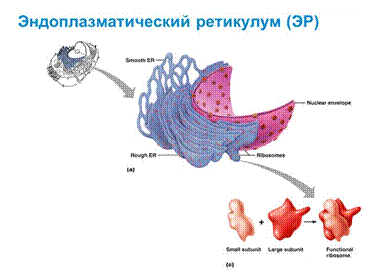
Гранулярная ЭПС содержит рибосомы на наружной стороне мембраны, обращенную к гиалоплазме. Гладкая ЭПС не содержит рибосомы. В скелетных мышцах ЭПС носит название саркоплазматический ретикулум. ЭПС пронизывает всю клетку. Полость ЭПС сообщается с перинуклеарным пространством ядра, а мембрана ЭПС с плазматической мембраной ядра. На рибосомах гранулярной ЭПС синтезируются секреторные белки, предназначенные для выведения из клетки, а также белки лизосом и внеклеточного матрикса.
Синтез указанных белков включает 5 этапов:
1. Связывание сигнал распознающий частицы (SRP), состоящий из 6-ти белков и малой цитоплазматической РНК (srpРНК) с сигнальной последовательность полипептидной цепи.
2. Взаимодействие комплекса матричной РНК (мРНК) – рибосома - SRP со специфическим белком-рецептором (SRP-рецептором), находящимся на цитоплазматической стороне мембраны.
3. Освобождение SRP от комплекса; связывание рибосомы с белок-транслоцирующим комплексом мембраны шероховатой ЭПС транслоконом и встраивание сигнальной последовательности в канал транслокона.
4. Возобновление трансляции и перенос растущей полипептидной цепи в полость ЭПС.
5. Удаление сигнальной последовательности от полипептида под действием фермента.
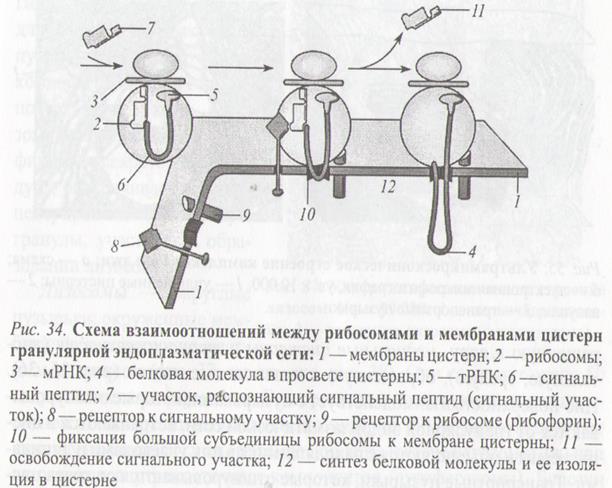
В полости ЭПС синтезированные водорастворимые белки подвергаются фолдингу и гликозилированию путём присоединения к белкам однотипных олигосахаридных цепочек. В результате фолдинга белки приобретают специфическую конформацию, и в составе транспортных пузырьков, отделяющихся от мембраны ЭПС, транспортируются в комплекс Гольджи, где подвергаются дальнейшим модификациям и сортировке.
Наряду с секреторными белками на гранулярной ЭПС синтезируется большая часть полуинтегральных и интегральных белков. В ЭПС происходит также синтез мембраны липидов и осуществляется «сборка» компонентов мембраны. При этом увеличения общей площади мембранной ЭПС не происходит. Это связано с тем, что некоторые фрагменты мембраны ЭПС постоянно утрачиваются в составе отделяющихся от нее транспортных пузырьков, которые переносят их в аппарат Гольджи и далее в другие мембранные структуры клетки:
- плазмалемму;
- лизосомы.
Кроме того, ЭПС, как считают, участвуют в образовании пероксисом. Таким образом, грЭПС служит «фабрикой» мембран для плазмалеммы, аппарата Гольджи, лизосом и других мембранных структур клетки. ЭПС обеспечивает также сегрегацию (разделение) белков, предназначенных на экспорт, и лизосомных гидролаз от литозольных белков.
Агранулярная (гладкая) эндоплазматическая сеть представляет собой замкнутую сеть трубочек, канальцев, цистерн и везикулярных образований размером 50 —100 нм и образуется, по-видимому, из гранулярной ЭПС.
На цитоплазматической поверхности гладкой ЭПС синтезируются жирные кислоты, холестерол и большая часть липидов клетки, в том числе почти все липиды, необходимые для построения клеточных мембран. Поэтому гладкую ЭПС нередко называют «фабрикой липидов». В клетках печени (гепатоцитах) с мембранами гладкого эндоплазматического ретикулума связан фермент, обеспечивающий образование глюкозы из глюкозо-6-фосфата. Эта реакция имеет большое значение в поддержании уровня глюкозы в организме человека. Кроме того, гладкая эндоплазматическая сеть осуществляет детоксикацию (обезвреживание) ксенобиотиков, а также ядовитых веществ, образующихся в процессе метаболизма и подлежащих удалению из организма. Большую роль в процессах детоксикации играют ферменты семейства цитохрома Р450, катализирующие реакции гидроксилирования или микросомального окисления. В результате указанных реакций липофильные (нерастворимые) вещества становятся более гидрофильными (растворимыми), заключаются в другие реакции и затем выводятся из организма.
Помимо указанных основных функций, гладкая эндоплазматическая сеть выполняет ряд дополнительных. В ее мембранах локализуются Са2+ - насосы, активно закачивающие ионы Са2+ из цитозоля в полость саркоплазматической сети. Поэтому концентрация ионов Са2+ в полости гладкой эндоплазматической сети достигает 10-3 моль/л, т.е. почти в 10 000 раз выше, чем в цитозоле. Под действием химических сигналов ионы Са2+ переносятся из полости гладкой ЭПС через ионные каналы, встроенные в ее мембрану, в гиалоплазму, где связываются с определенными белками, влияющими на множество внутриклеточных процессов: активацию или инактивацию ферментов, экспрессию генов, освобождение антител из клеток иммунной системы и др.
В мышечных волокнах гладкая ЭПС имеет структурные и функциональные особенности и называется саркоплазматической сетью. При возбуждении плазмалеммы выход ионов Са2 из гладкой эндоплазматической сети в цитозоль стимулирует сокращение миофибрилл.
В организме человека эндоплазматическая сеть особенно хорошо развита в клетках, синтезирующих гормоны, в клетках печени (гепатоцитах) и некоторых клетках почек.
Комплекс Гольджи (КГ), или аппарат Гольджи, - пластинчатый комплекс, расположен вблизи ядра, между ЭПС и плазмалеммой. Его структурно-функциональная единица — диктиосома представляет собой стопку из 5—20 плоских одномембранных мешочков (цистерн), имеющих диаметр около 1 мкм, внутренние полости которых не сообщаются друг с другом. Количество таких мешочков в стопке обычно не превышает 5-10, а расстояние между ними составляет 20—25 нм.
В каждой диктиосоме различают три части, проксималъную (cis-полюс), обращенную к ЭПС, медиальную и дистальную (trans- полюс), обращенную к плазмалемме. К дистальной цистерне транс-полюса примыкают многочисленные трубочки и пузырьки, образующие транс-сеть аппарата Гольджи.
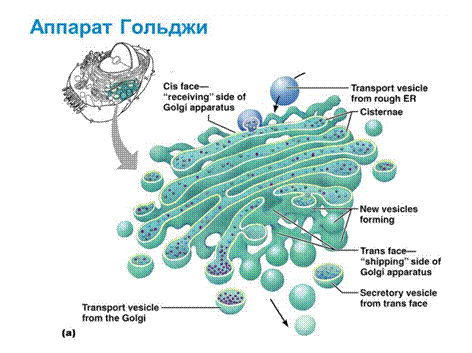
Белки, синтезированные на шероховатой эндоплазматической сети в составе отпочковывающихся от нее мембранных пузырьков, покрытых клатрином, транспортируются к цис-полюсу аппарата Гольджи. В аппарате Гольджи осуществляется химическая модификация транспортируемых белков. Этот процесс происходит поэтапно по мере транспорта белков от цис- к транс-полюсу аппарата Гольджи, каждая цистерна которого содержит характерный для нее набор ферментов. Гликопротеины, несущие первоначально одинаковые олигосахаридные цепочки, в проксимальных цистернах подвергаются последовательной перестройке, специфической для белков каждого вида. В процессе гликозилирования одни олигосахаридные остатки удаляются, другие добавляются с образованием дополнительных ветвей олигосахаридов. В аппарате Гольджи некоторых специализированных клеток синтезируются полисахариды, которые, соединяясь с белками, образуют протеогликаны, составляющие основу вещества межклеточного матрикса. Таким образом, химическая перестройка белков в цистернах цис-полюса аппарата Гольджи осуществляется главным образом путем их гликозилирования, тогда как в транс-полюсе происходит в основном сульфатирование и фосфорилирование белков. Некоторые белки в комплексе Гольджи подвергаются протеолитическому расщеплению, в результате чего первоначально неактивные молекулы приобретают биологическую активность. Например, таким путем в клетках поджелудочной железы образуется инсулин.
В комплексе Гольджи протекают реакции синтеза мембранных гликолипидов. При этом углеводные компоненты в молекулах синтезированных мембранных гликолипидов, также как и у гликопротеинов, всегда оказываются обращенными в полость цистерн аппарата Гольджи. Этим объясняется топология углеводных остатков молекул мембранных гликолипидов и гликопротеинов при включении их в состав плазмалеммы, где они участвуют образовании гликокаликса.
В транс-сети осуществляется сортировка белков и упаковка их в мембранные пузырьки, покрытые клатрином. Белки, предназначенные для экспорта, заключаются в одни пузырьки, лизосомные белки — в другие, мембранные белки — в третьи. В специальные пузырьки упаковываются также белки, присущие ЭПС, например ферменты фолдинга, которые случайно оказались в комплексе Гольджи.
Сортировка белков осуществляется при помощи встроенных в мембрану транс-сети КГ особых белков-рецепторов, способных специфически взаимодействовать с определенными химическими группировками — маркерами молекул отбираемых белков. В результате участок мембраны, несущей рецепторы, связанные с маркерами отбираемых молекул, обособляется от КГ с образованием мембранного пузырька, покрытого клатрином, транспортирующего нужное вещество к месту назначения. Например, маркером лизосомальных ферментов служит присоединяющийся к ним в комплексе Гольджи олигосахарид, содержащий остаток маннозо-6-фосфата. Для белков ЭПС — ферментов фолдинга и др., которые случайно оказались в КГ, таким маркером служит последовательность из четырех аминокислот, по которой они узнаются и упаковываются в пузырьки, а затем возвращаются обратно в ЭПС. Этот процесс получил название рециклизации.
Секреция экспортных белков осуществляется посредством конститутивной и регулируемой секреции.
В случае конститутивной секреции, которая характерна для всех клеток, транспортные пузырьки непрерывно переносятся от аппарата Гольджи к плазмалемме. Таким путем, например, доставляются к поверхности клетки элементы гликокаликса. А также многие продукты, составляющие основу межклеточного вещества. Посредством конститутивной секреции осуществляется также транспорт мембранных белков плазмалеммы.
Регулируемая секреция свойственна лишь специализированным клеткам, секретирующим биологически активные вещества в ответ на действие сигналов, например гормонов.
Таким образом, основными функциями комплекса Гольджи являются химическая модификация, накопление, сортировка, упаковка в секреторные пузырьки и транспорт по назначению белков и липидов, синтезированных в эндоплазматическом ретикулуме. В комплексе Гольджи образуются лизосомы и синтезируются некоторые полисахариды.
Дата добавления: 2017-10-04; просмотров: 3287;
Поиск по сайту
Узнать еще
- Двумембранные органоиды. Митохондрии.
- История создания клеточной теории. Клеточная оболочка. Немембранные органоиды. Включения
- Лекция. Одномембранные органоиды
- Мембранные органоиды растительной клетки
- Митохондрии, пластиды, рибосомы и другие органоиды.
- Немембранные органеллы
- Немембранные органеллы общего назначения.
- НЕМЕМБРАННЫЕ ОРГАНЕЛЛЫ. ЖГУТИКИ И УНДУЛИПОДИИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
