Двумембранные органоиды. Митохондрии.
Двумембранные, полуавтономные органоиды, которые обеспечивают клетку основной энергией, получаемой в результате окисления органических молекул с помощью кислорода. Присутствуют в клетках грибов, растений и животных. Форма, размеры и количество митохондрий чрезвычайно варьируют. По форме митохондрии могут быть палочковидными, округлыми, спиральными, чашевидными, разветвленными. Длина митохондрий колеблется в пределах от 1,5 до 10 мкм, диаметр – от 0,25 до 1,00 мкм. Количество митохондрий в клетке может достигать 500-1000 в тех клетках, которым нужно очень много энергии, т.е. количество митохондрий зависит от метаболической активности клетки. Кроме того, наибольшее количество митохондрий находится в участках клетки, которые потребляют больше энергии – вблизи ионных насосов, в мышечных клетках – вблизи миофибрилл.
|
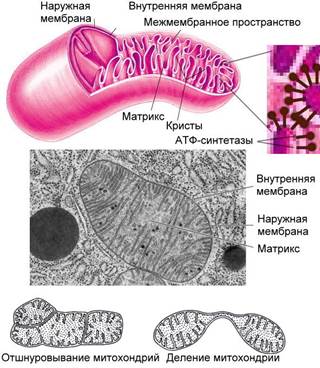 Рис. . Строение и образование митохондрий
Рис. . Строение и образование митохондрий
Ширина межмембранного пространства (протонного резервуара) – 10-20 нм.
Внутреннее пространство митохондрий заполнено внутренней средой, матриксом. В матриксе содержатся различные ферменты (например, ферменты цикла Кребса), кольцевые ДНК, содержащие всего 37 генов в количестве от 1 до 50 таких молекул и собственный белоксинтезирующий аппарат.
Митохондриальные ДНК не связаны с белками («голые»), прикреплены к внутренней мембране. Для построения митохондрии требуется гораздо больше белков, чем кодирует кольцевая ДНК митохондрий, поэтому следует отметить, что информация о большинстве митохондриальных белков содержится в ядерной ДНК, и эти белки синтезируются в цитоплазме клетки, а затем транспортируются в митохондрии. Митохондриальная ДНК кодирует иРНК, тРНК, рРНК, формируя собственные системы репликации ДНК, транскрипции и трансляции некоторых белков. Генетический код ДНК митохондрий имеет несколько отличий от генетического кода эукариот. Рибосомы митохондрий прокариотического типа (70S-типа), большая часть рибосомальных белков синтезируется в цитоплазме, а затем транспортируется в митохондрии.
Размножение. Митохондрии живут около 10 суток, способны размножаться путем деления или отшнуровывания новых митохондрий от ранее существующих; разрушение их происходит с помощью автофагии. Наследуются митохондрии у многих видов, в том числе и у человека, по материнской линии, митохондрии отца разрушаются в процессе оплодотворения.
Происхождение. Согласно теории симбиогенеза митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина (анаэробную архебактерию) образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения, как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу – 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками. Но в процессе эволюции большая часть генов митохондрий переместилась в ядро, и синтез большинства митохондриальных белков происходит в цитоплазме клетки, т.е. митохондрии являются полуавтономными органоидами эукариотической клетки.
Функции. Основная функция – окисление органических молекул с образованием энергии в форме тепла и в форме АТФ. Но в клетках бурого жира (например у медведя во время зимнего сна) митохондрии не образуют АТФ, вся энергия выделяется в форме тепла и поддерживает определенную температуру тела. Кроме этого в митохондриях происходит синтез некоторых (около 5%) митохондриальных белков.
Пластидыхарактерны только для растительных клеток. Различают три основных типа пластид: лейкопласты – бесцветные пластиды в клетках неокрашенных частей растений, хромопласты – окрашенные пластиды обычно желтого, красного и оранжевого цвета, хлоропласты – зеленые пластиды.
|
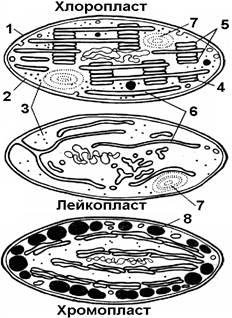 Рис. . Строение пластид
1 – наружная мембрана; 2 – внутренняя мембрана; 3 –строма; 4 – тилакоид;5- грана; 6 – ламелла; 7 – зерно первичного крахмала; 8 – липидные капли.
Рис. . Строение пластид
1 – наружная мембрана; 2 – внутренняя мембрана; 3 –строма; 4 – тилакоид;5- грана; 6 – ламелла; 7 – зерно первичного крахмала; 8 – липидные капли.
Хлоропласты содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Основная функция хлоропластов – фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Функция: синтез, накопление и хранение запасных питательных веществ. В зависимости от накопленных органических веществ различают амилопласты – лейкопласты, которые синтезируют и накапливают крахмал, элайопласты – масла, протеинопласты – белки. Кроме того, в одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты – каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различна: в виде кристаллов, липидных капель и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко – корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды – мелкие органоиды, содержащиеся в образовательных тканях зародыша семени. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету) и хромопласты (при созревании корнеплодов). Хлоропласты в темноте становятся лейкопластами, осенью, при разрушении хлорофилла – хромопластами (изменение окраски листьев связано с выявлением каротиноидов – пигментов желтого цвета ксантофиллов и оранжевых пигментов каротинов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Строение и функции ядра. Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных). Форма ядра обычно сферическая, диаметр от 3 до 10 мкм.
|
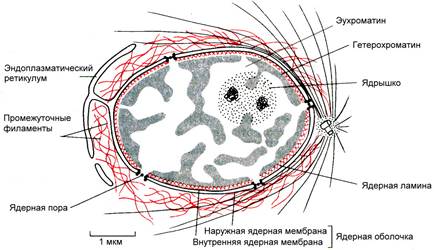 Рис. . Строения ядра
Рис. . Строения ядра
Внутренняя мембрана гладкая, под ней находится ядерная ламина – часть кариоскелета, состоящая из промежуточных филаментов. Ядерная ламина поддерживает форму ядра, участвует в упорядоченной укладке хроматина внутри ядра и организации ядерных пор.
Кариоплазма (ядерный сок, нуклеоплазма) – внутреннее среда ядра, в которой располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды.
Ядрышко представляет собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и варьирует от 1 до 10 и более. Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают. Ядрышко образуется на определенных участках хромосом, несущих информацию о структуре рРНК. Такие участки называются ядрышковыми организаторами и содержат многочисленные копии генов, кодирующих рРНК. Из рРНК и белков, поступающих из цитоплазмы, формируются субъединицы рибосом. Таким образом, ядрышко представляет собой скопление рРНК и рибосомальных субъединиц на разных этапах их формирования.
Хроматин – внутренние нуклеопротеидные структуры ядра, окрашивающиеся некоторыми красителями и отличающиеся по форме от ядрышка. Хроматин имеет вид глыбок, гранул и нитей. Химический состав хроматина: ДНК (30-45%), гистоновые белки (30-50%), негистоновые белки (4-33%), т.о. хроматин является дезоксирибонуклеопротеидным комплексом (ДНП). В зависимости от функционального состояния хроматина различают: гетерохроматин и эухроматин. Эухроматин – генетически активные, гетерохроматин – генетически неактивные участки хроматина. Эухроматин при световой микроскопии не различим, слабо окрашивается и представляет собой деконденсированные (деспирализованные, раскрученные) участки хроматина. Гетерохроматин под световым микроскопом имеет вид глыбок или гранул, интенсивно окрашивается и представляет собой конденсированные (спирализованные, уплотненные) участки хроматина. Хроматин – форма существования генетического материала в интерфазных клетках. Во время деления клетки (митоз, мейоз) хроматин преобразуется в хромосомы.
Функции ядра: хранение наследственной информации и передача ее дочерним клеткам в процессе деления, регуляция жизнедеятельности клетки путем регуляции синтеза различных белков, место образования субъединиц рибосом.
|
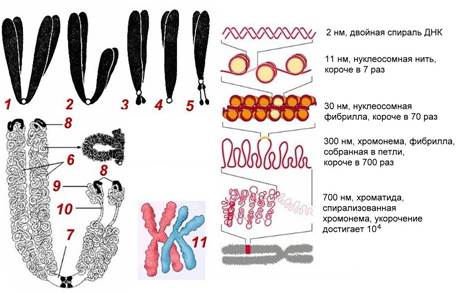 Рис. . Строение хромосом. Компактизация ДНК
1 – равноплечая (метацентрическая) хромосома; 2 – неравноплечая (субметацентрическая) хромосома; 3 – резко неравноплечая (акроцентрическая) хромосома; 4 – одноплечая (телоцентрическая) хромосома; 5 – спутничная хромосома; 6 – хроматиды; 7 – центромера; 8 – теломеры; 9 – спутники; 10 – ядрышковые организаторы; 11 – гомологичные хромосомы.
Рис. . Строение хромосом. Компактизация ДНК
1 – равноплечая (метацентрическая) хромосома; 2 – неравноплечая (субметацентрическая) хромосома; 3 – резко неравноплечая (акроцентрическая) хромосома; 4 – одноплечая (телоцентрическая) хромосома; 5 – спутничная хромосома; 6 – хроматиды; 7 – центромера; 8 – теломеры; 9 – спутники; 10 – ядрышковые организаторы; 11 – гомологичные хромосомы.
длина ДНК одной хроматиды может достигать нескольких сантиметров. Понятно, что молекула такой длины не может располагаться в клетке в вытянутом виде, а подвергается укладке, приобретая определенную трехмерную структуру, или конформацию. Выделяют следующие уровни пространственной укладки ДНК и ДНП: нуклеосомный (накручивание ДНК на белковые глобулы – нуклеосомы). Каждая глобула, нуклеосома состоит из 8 гистоновых белковых молекул, ДНК делает вокруг нуклеосомы 1,75 оборота. Нуклеосомы спирально закручиваются, образуя нуклеосомную фибриллу; нуклеосомная фибрилла собирается в крупные сближенные петли, образуя хромонему, хромонема закручивается в суперспираль, образуя хроматиду. Хромосома перед делением клетки состоит из двух хроматид. В хромосоме различают первичную перетяжку, плечи хромосомы (части хромосомы по обе стороны от первичной перетяжки), теломеры (концевые участки плеч, защищающие хромосомы от слипания). Некоторые хромосомы имеют вторичные перетяжки, отделяющие часть хромосомы, называемую спутником (у человека пять пар хромосом имеют вторичные перетяжки). В области вторичных перетяжек копии генов, несущих информацию о строении рРНК, поэтому эти хромосомы называются ядрышкообразующими.По положению центромеры хромосомы делят на метацентрические (равноплечие), субметацентрические (неравноплечие), акроцентрические (резко неравноплечие), телоцентрические (одноплечие) и спутничные.
|
 Рис. . Идиограмма кариотипа человека
Рис. . Идиограмма кариотипа человека
Соматические клетки содержат диплоидный, двойной – 2n набор хромосом. Половые клетки –гаплоидный, одинарный – n. Диплоидный набор дрозофилы – 8, шимпанзе – 48, речного рака – 196 хромосом. Хромосомы диплоидного набора разбиваются на пары, хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
Кариотип – совокупность признаков о числе, размерах и строении метафазных хромосом, характерных для вида.
Идиограмма– графическое изображение кариотипа. У представителей разных видов кариотипы разные, одного вида – одинаковые. Аутосомы – хромосомы, одинаковые для мужского и женского кариотипов. Половые хромосомы – хромосомы, по которым мужской кариотип отличается от женского. Кариотип человека (2n = 46, n = 23) содержит 22 пары аутосом и 1 пару половых хромосом. Половые хромосомы женщины – «ХХ», мужчины – «ХУ». Х-хромосома – субметацентрическая, У-хромосома – акроцентрическая.
Функции хромосом – хранение наследственной информации и передача генетического материала от материнской клетки к дочерним.
1.
| <== предыдущая лекция | | | следующая лекция ==> |
| Лекция. Одномембранные органоиды | | | Прокариоты. Описание. Характеристика |
Дата добавления: 2017-11-21; просмотров: 5491;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
