Модель. Репликация ДНК
Гены и хромосомы
Джеймс Уотсон и Фрэнсис Крик не только постулировали структуру ДНК, но и объяснили, каким образом может передаваться генетическая информация. Это происходит в три этапа:
- репликация (копирование родительской ДНК с образованием дочерних ДНК);
- транскрипция (переписывание генетической информации в форме РНК);
- трансляция (перевод информации с РНК на белковую форму).
Роль ДНК как носителя генетической информации подтверждается опытными фактами. Так, Освальдом Эвери, Колином Мак-Леодом и Маклином Мак-Карти было показано, что ДНК, выделенная из одного штамма бактерий, способна перейти в клетки другого штамма и трансформировать их, передавая некоторые наследственные признаки донора.
Ряд вирусов имеет одноцепочечную молекулу ДНК, но у большинства ДНК-содержащих вирусов ДНК двухцепочечная, и она линейна или замкнута в кольцо. В клеточных организмах ДНК содержится в хромосомах. Бактериальная хромосома содержит гораздо большую по размерам молекулу ДНК, также свёрнутую в кольцо. Эти кольца сверхспирализированы: двойная спираль, прежде чем её концы были соединены, была частично раскручена. Такой эффект позволяет молекуле разместиться более компактно.
Хромосомы эукариот представляют собой линейную молекулу ДНК. Эукариотическая ДНК обматывает белковые частицы – гистоны, располагающиеся вдоль ДНК через определённые интервалы, образуя хроматин – волокна, из которых состоят хромосомы. Комплексы участков ДНК и гистонов называются нуклеосомами. Нуклеосомы упорядочены в пространстве, за счёт чего достигается плотная упаковка ДНК в хромосоме.
Размеры ДНК зависят от типа организма. Физическая длина ДНК вирусов составляет десятки микрометров, бактерий – миллиметры, а человека – 2 метра. Общая длина всех ДНК человека составляет 2∙1010 км.
Структурными генами называются участки ДНК, кодирующие белковые цепи, т-РНК и р-РНК. В классической биологии гены определяли как часть хромосомы, определяющей какой-то наследственный признак. Более точное определение дала молекулярная биология: ген – фрагмент генетического кода, который может синтезировать только один определённый полипептид или РНК. Наряду со структурными генами ДНК содержит регуляторные последовательности. Они могут обозначать начало или конец структурного гена, запускают или прекращают транскрипцию.
Ген среднего полипептида имеет длину в десятые доли микрометра. Гены, кодирующие т-РНК, значительно короче. Вирусные ДНК содержат небольшое количество генов, бактериальные – уже тысячи генов. Бактерии защищают собственную ДНК метилированием некоторых оснований. Чужеродная ДНК, не имеющая строго определённых опознавательных метильных групп, разрушается эндонуклеазами.
В генах эукариот содержатся нетранслирующиеся вставки – интроны. Их значение до конца не понятно; возможно, они делят гены на отдельные участки, которые могут рекомбинировать в ходе эволюции с образованием новых генов. Вообще, нетранслируемого материала в клетках эукариот довольно много – возможно, около 10 %.
Часть генов «работает» только при определённых условиях. Так, ген, регулирующий синтез инсулина, способен выполнять свои функции только в специальных клетках поджелудочной железы, а гемоглобин вырабатывается только в том случае, если гены, отвечающие за его синтез, находятся в клетках молодых эритроцитов.
Некоторые гены повторяются в ДНК во множестве копий. Таковыми, например, являются гены, синтезирующие кератин перьев цыплят, – для быстрого роста животного необходима большая скорость производства «строительного материала», и один ген с этой работой не справлялся бы.
| Клетки организма данного вида (даже принадлежащие разным тканям) содержат ДНК с одинаковым нуклеотидным составом, и этот состав не зависит ни от питания, ни от окружающей среды, ни от возраста организма. Нуклеотидный состав ДНК разных видов различен. |
Доказать, что проявление определённого признака, унаследованного организмом, является результатом действия конкретного гена, достаточно трудно. Как правило, признаки проявляются в результате взаимодействия нескольких генов, либо один ген влияет на наследование нескольких признаков сразу. Например, наличие окраски у многих растений зависит только от того, окажутся ли рядом два необходимых гена. Один-единственный ген может вызвать у животных изменение целого ряда систем.
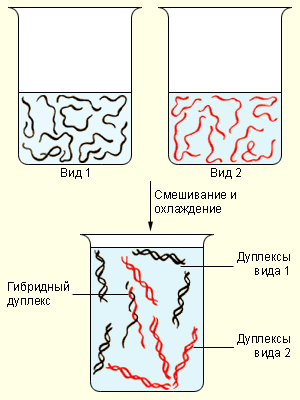
|
| Гибридизация ДНК |
При нагревании или других экстремальных условиях спираль ДНК расплетается, и её цепи разделяются. Если этот процесс не дошел до конца, то он легко обратим вспять. При полной денатурации обратный процесс будет протекать значительно медленнее: комплементарным участкам цепей сначала нужно «отыскать» друг друга, после чего две цепи сравнительно быстро «застёгиваются» наподобие молнии, вновь образуя двойную спираль. Одноцепочечная ДНК одного вида организма может образовать гибрид с ДНК другого вида, при условии, что нуклеотидные последовательности цепей хоть немного похожи. Естественно, спариваются только отдельные участки цепей, которые комплементарны друг другу. Исследуя образование таких гибридов, можно сделать вывод об отноРепликация и транскрипция ДНК
При синтезе «неинформационной» молекулы (например, гликогена) чистота конечного продукта обеспечивается специальным ферментом. Для фермента характерна субстратная специфичность, то есть его активный центр способен присоединять только молекулу UDP-глюкозы и нередуцирующий конец молекулы гликогена, которая должна быть удлинена. Таким образом, активный центр фермента можно рассматривать как «матрицу», поскольку между молекулами субстрата осуществляется комплементарная подгонка.
При синтезе макромолекул ДНК, РНК или белков один активный центр фермента не в состоянии обеспечить специфическую последовательность четырёх кодирующих единиц. Он может связывать между собой только один или несколько «строительных блоков», а нуклеиновые кислоты содержат в своём составе тысячи нуклеотидов. Поэтому природа пошла здесь по другому пути: матрицей для синтеза цепи молекулы ДНК служит другая цепь ДНК.
Транскрипция ДНК в ходе деления клеток начинается с разделения двух цепей, каждая из которых становится матрицей, синтезирующей нуклеотидную последовательность новых цепей. Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований. Репликация катализуется несколькими ДНК-полимеразами, а транскрипция – ферментом РНК-полимеразой. После репликации дочерние спирали закручиваются обратно уже без затрат энергии и каких-либо ферментов.
Сравнительно неплохо изучен процесс репликации и транскрипции ДНК бактерий. Их ДНК способна реплицироваться, не распрямляясь в линейную молекулу, то есть в кольцевой форме. Процесс, по-видимому, начинается на определённом участке кольца и идёт сразу в двух направлениях (в одном – непрерывно, во втором – фрагментарно с последующим «склеиванием» фрагментов). Инициация репликации находится под контролем клеточной регуляции. Скорость репликации ДНК составляет около 45 000 нуклеотидов в минуту; таким образом, родительская вилка расплетается со скоростью 4500 об/мин.
Частота ошибок при ДНК-репликации не превышает 1 на 109–1010 нуклеотидов. Столь высокая степень точности воспроизведения информации определяется не только комплементарностью нуклеотидов, но и действием ДНК-полимераз, которые способны распознать ошибку в образующемся коде и исправить её. Следует заметить, что точность воспроизведения РНК и белков в тысячи раз ниже. Это связано с тем, что транскрипция и трансляция, затрагивающие только одну клетку, – не столь жизненно важные процессы, как репликация, которая определяет будущее всего вида.
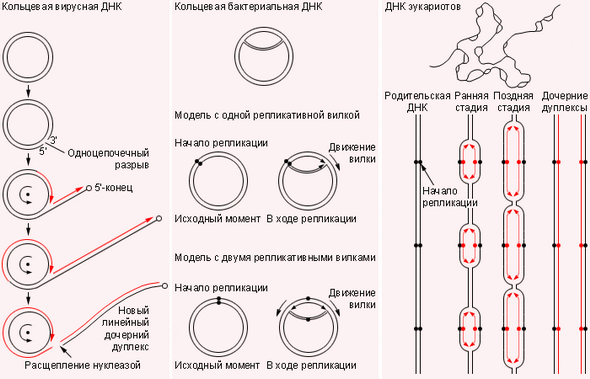
|
| Репликация ДНК прокариот и эукариот |
Репликация эукариот при такой же схеме длилась бы несколько месяцев (скорость движения репликативных вилок составляет всего микрометр в минуту). Поэтому в ДНК эукариот процесс начинается одновременно в сотнях и тысячах точек. Все хромосомы в клетке должны реплицироваться одновременно, и одновременно в клетке работают многие тысячи вилок.
Между репликацией и транскрипцией есть существенная разница: в первом случае копируется вся молекула ДНК, во втором, как правило, только отдельные гены. Минимальная длина и-РНК определяется длиной полипептидной цепи, для которой она предназначена. В идентификации последовательностей нуклеотидов, обозначающих начало и конец синтезирующих РНК генов, ещё много неясного.
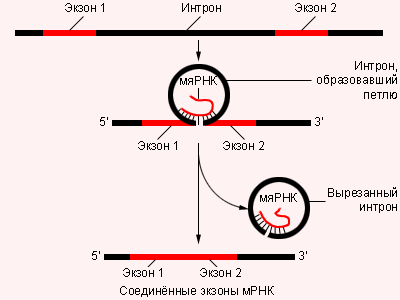
|
| Роль малой ядерной РНК в процессе транскрипции |
Молекулы р-РНК и т-РНК образуются из более длинных предшественников – гетерогенных ядерных РНК (гя-РНК). Длина гя-РНК увеличена за счет нетранслирующихся интронов, которых в конечных РНК уже нет. Интроны удаляются при помощи малой ядерной РНК. мя-РНК комплементарна нуклеотидам на концах интронов – она временно соединяется с ними, стягивая интрон в петлю. Концы кодирующих фрагментов соединяются, после чего интрон благополучно удаляется из цепи.
Некоторые РНК-содержащие вирусы животных при помощи РНК-зависимой ДНК-полимеразы способны синтезировать ДНК, комплементарную по отношению к вирусной РНК. Она встраивается в геном эукариотической клетки, где может многие поколения оставаться в скрытом состоянии. При определённых условиях (например, воздействии канцерогенов) вирусные гены могут активироваться, и здоровые клетки превратятся в раковые.
Модель. Репликация ДНК
сительном родстве разных видов.
Синтез белка
Синтез белка (трансляция) является самым сложным из биосинтетических процессов: он требует очень большого количества ферментов и других специфических макромолекул, общее количество которых, видимо, доходит до трёхсот. Часть из них к тому же объединены в сложную трёхмерную структуру рибосом. Но несмотря на большую сложность синтез протекает с чрезвычайно высокой скоростью (десятки аминокислотных остатков в секунду). Процесс может замедляться и даже останавливаться ингибиторами-антибиотиками.
В пятидесятых годах XX века было установлено, что синтез белка происходит в рибонуклеопротеиновых частицах, называющихся рибосомами. Диаметр рибосомы бактерии E. coli составляет 18 нм, а их общее количество – десятки тысяч в клетке. Рибосомы эукариот несколько крупнее (21 нм). Сам процесс протекает в пять этапов.
1.Активация аминокислот. Каждая из 20 аминокислот белка соединяется ковалентными связями к определённой т-РНК, используя энергию АТФ. Реакция катализуется специализированными ферментами, требующими присутствия ионов магния.
2.Инициация белковой цепи. и-РНК, содержащая информацию о данном белке, связывается с малой частицей рибосомы и с инициирующей аминокислотой, прикреплённой к соответствующей т-РНК. т-РНК комплементарна с находящимся в составе и-РНК триплетом, сигнализирующим о начале белковой цепи.
3.Элонгация. Полипептидная цепь удлиняется за счёт последовательного присоединения аминокислот, каждая из которых доставляется к рибосоме и встраивается в определённое положение при помощи соответствующей т-РНК. В настоящее время генетический код полностью расшифрован, то есть всем аминокислотам поставлены в соответствие триплеты нуклеотидов. Элонгация осуществляется при помощи белков цитозоля (так называемые факторы элонгации).
4.Терминация. После завершения синтеза цепи, о чём сигнализирует ещё один специальный кодон и-РНК, полипептид высвобождается из рибосомы.
5.Сворачивание и процессинг. Чтобы принять обычную форму, белок должен свернуться, образуя при этом определённую пространственную конфигурацию. До или после сворачивания полипептид может претерпевать процессинг, осуществляющийся ферментами и заключающийся в удалении лишних аминокислот, присоединении фосфатных, метильных и других групп и т. п.
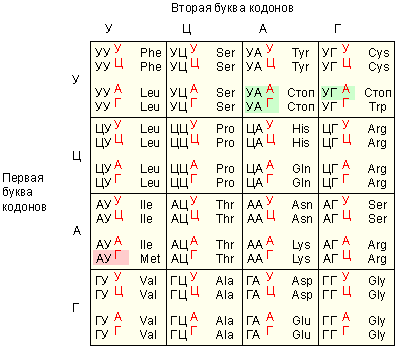
|
| Генетический код |
Генетический код обладает рядом особенностей. Во-первых, в коде отсутствуют «знаки препинания», то есть сигналы, показывающие начало и конец кодонов. Во-вторых, 3 нуклеотидных триплета (УАГ, УАА, УГА) не соответствуют никакой аминокислоте, а обозначают конец полипептидной цепи, а кодон АУГ сигнализирует о начале цепи либо (если он в середине последовательности) об аминокислоте метионине. Многие аминокислоты могут кодироваться несколькими различными кодонами. Все кодоны аминокислот одинаковы у всех изученных организмов: от вируса до человека. Создаётся впечатление, что все организмы на Земле происходят от единого генетического предка. Впрочем, в последнее время в митохондриях клеток человека были обнаружены кодоны, не совпадающие с «нормальным» словарём. Их наличие представляет собой загадку для ученых.
Синтез белка требует больших затрат энергии – 24,2 ккал/моль. После окончания синтеза белок при помощи специального полипептидного лидера доставляется к месту своего назначения.
Синтез белка контролируют гены-операторы. Совокупность рабочих генов – операторов и структурных генов – называется оперон. Опероны не являются самостоятельной системой, а «подчиняются» генам-регуляторам, отвечающим за начало или прекращение работы оперона. Свой контроль гены-регуляторы осуществляют при помощи специального вещества, которое они при необходимости синтезируют. Это вещество реагирует с оператором и блокирует его, что влечёт за собой прекращение работы оперона. Если же вещество реагирует с небольшими молекулами – индукторами, это будет являться сигналом к возобновлению работы системы.
Модель оперонов была разработана на микроорганизмах, но она соответствует и принципу работы генома эукариот. У последних гены образуют сложные системы, называемые супергенами, которые могут одновременно кодировать множество идентичных друг другу молекул белка.
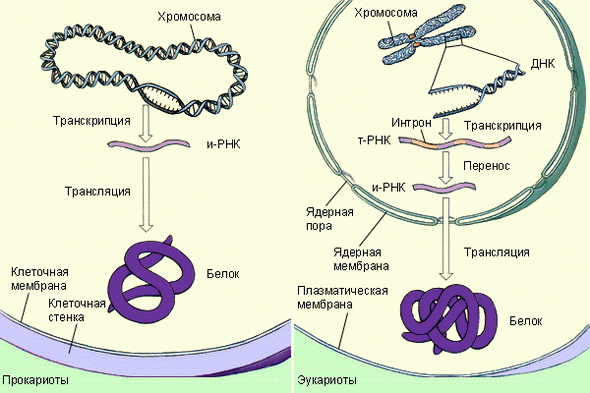
|
| Синтез белка у прокариот и эукариот |
Все многоклеточные организмы развиваются из одной-единственной клетки – зиготы. Процесс дифференцировки клеток, видимо, связан с управлением синтезом белка генами-регуляторами, но каким конкретно образом осуществляется это управление – пока остаётся неясным.
Репликация ДНК
|
| Модель. Репликация ДНК |
Репликация ДНК – это процесс копирования дезоксирибонуклеиновой кислоты, который происходит в процессе деления клетки. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация катализируется несколькими ДНК-полимеразами. После репликации дочерние спирали закручиваются обратно уже без затрат энергии и каких-либо ферментов.
Скорость репликации составляет порядка 45 000 нуклеотидов в минуту, а родительская вилка вращается со скоростью 4500 об/мин. Частота ошибок при репликации не превышает 1 на 109–1010 нуклеотидов. ДНК эукариот с такой скоростью реплицировалась бы несколько месяцев, поэтому в хромосомах ядерных клеток репликация производится сразу в сотнях и тысячах точек.
Анимация модели управляется кнопками «Старт»/«Стоп» и «Сброс».
| <== предыдущая лекция | | | следующая лекция ==> |
| Оптимальные условия хранения неснятой и снятой кинопленки. | | | Герметизация фиссур |
Дата добавления: 2019-12-09; просмотров: 742;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
