СТРОЕНИЕ БРЮШНОГО СЕГМЕНТА И ЕГО МУСКУЛАТУРА
Тергит и стернит брюшного сегмента соединены более или менее обширно мембраной. В мембране расположено дыхальце (рис. 223 А). Однако при разрастании тергита дыхальце может оказаться на этом последнем (рис.223 С). Вопрос о плейритах довольно сложен. Плейриты считаются производными конечностей (стр.160), но так как брюшные сегменты по большей части не им ют конечностей, то и наличие плейритов становится неясным. Между тергитом и стернитом нередко имеются склериты, иногда их несколько (рис. 222 В). Их называют латеротергитами или латеростернитами смотря по их расположению — ближе к тергитам или стернитам. Происхождение их неясно.
Боковые границы спинной области в брюшке определяются продолжением линий, которые в груди идут над и под плейритами. У гусениц отыскать ее нередко помогает расположение волосков. Если на брюшке есть конечности, то о брюшных плейритах можно говорить более определенно.
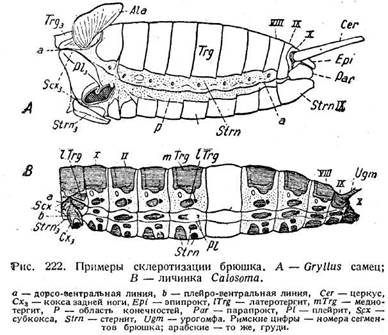
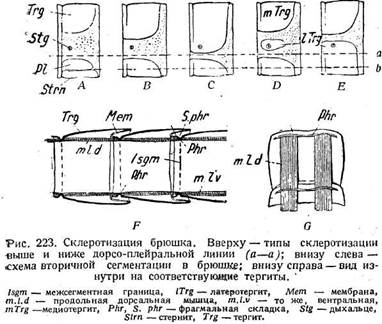
Распределение склеритов в общем такое же, как в груди. Первичные сочленовные промежутки склеротизируются и утрачивают подвижность, а к их фрагмальным складкам прикрепляются продольные мышцы (рис. 223). Вторичная сочленовная мембрана образуется в задней части сегмента. Благодаря своему впячиванию она спрятана под склеритом. В отличие от груди передние склериты одинаково надвигаются на задние как на спинной, так и на брюшной сторонах.
Относительная степень развития тергита, стернита и плейральной мембраны в брюшном сегменте весьма различна (рис. 224). Кузнечики с их обширной плейральной областью наиболее близки к основной схеме. У некоторых жуков тергит плоский, плейральная мембрана смещена дорсально, а стернит выпуклый.
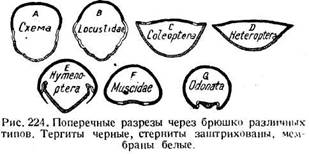
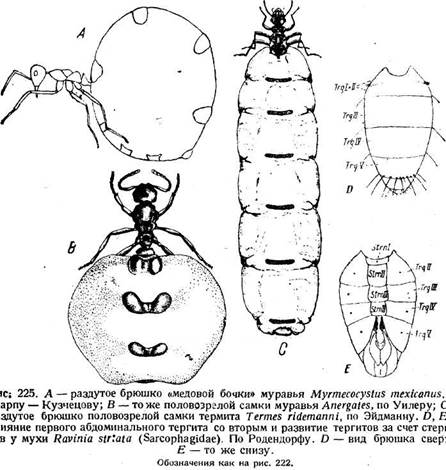
Сходное состояние характерно для клопов, хотя плейральная область у них не подвергается дорсальному смещению. У перепончатокрылых выпуклый тергит надвигается на выпуклый стернит, а плейральная мембрана спрятана между ними. У мух и стрекоз тергит брюшка получает сильное преобладание над стернитом (рис. 225). Иногда брюшко сильно растягивается за счет сочленовных мембран. Уже упоминались самки термитов и некоторые муравьи, у которых, вследствие колоссального развития яичника, брюшко увеличивается в объеме во много раз, так что большую часть его поверхности составляют сочленовные мембраны, на которых тергиты, стерниты и дыхальца имеют вид небольших пятнышек.
МУСКУЛАТУРА БРЮШКА
Брюшные сегменты имеют пять основных групп мышц: спинные продольные, брюшные продольные, дорсовентральные, поперечные и дыхальцевые.
Две первых группы продольных мышц близко сходны с тем, что мы видели в груди, т. е. они тянутся от одной фрагмы к другой, причем

одна группа лежит на спинной (рис. 223), другая — на брюшной стороне. Каждая из них может разделяться на несколько отдельных мышц, которые лежат одна над другой у стенки тела (рис. 226 Л). При этом одни продольные мышцы оказываются внутренними, другие наружными. Внутренние мышцы, притягивая друг к другу фрагмальные складки, сближают сегменты и укорачивают брюшко. При неодинаковом сокращении справа и слева они вызывают изгибание тела; Наружные продольные мышцы короче внутренних. Каждая наружная мышца одним концом прикреплена к переднему краю вторичного сегмента, а другим либо к средине сегмента (рис. 226 С), либо неподалеку от его заднего края, направляясь таким образом назад (рис. 226 D I. d. e). В первом случае ее действие аналогично таковому внутренней продольной мышцы,, т. е. она надвигает один сегмент на другой. Во втором —наружная мышца
«служит антагонистом внутренней, ибо она выдвигает край заднего сегмента .из-под переднего. Так называемые парадорсальные мышцы (рис. 226 В, pd) проходят между дорсо-вентральными (dv) и стенкой тела.
Дорсо-вентральные мышцы, если и гомологичны грудным дорсо-вентральным, то отличаются от них очень сильно (рис. 226 A, dv). Они лежат латерально и идут либо от тергита к стерниту (t. strn), либо от тергита в плейральную область (t. pi), либо соединяют последнюю со стер-нитом (strn. pi). Во всех этих случаях они сплющивают брюшко, что важно для дыхательных движений. Но если тергит заходит нижним краем на стернит, то отдельные группы дорсо-вентральных мышц могут быть антагонистами продольных мышц и расширять брюшко при своем сокращении (рис. 226 #, dilat). Действие их различно в зависимости от особенности склеро-"тизации. Если тергит мягок, а стернит склеротизирован, то при сокращении этих мышц движется тергит, при обратном соотношении движется стернит.
Если расширители брюшка отсутствуют, то их действие заменяется эластичностью хитина.
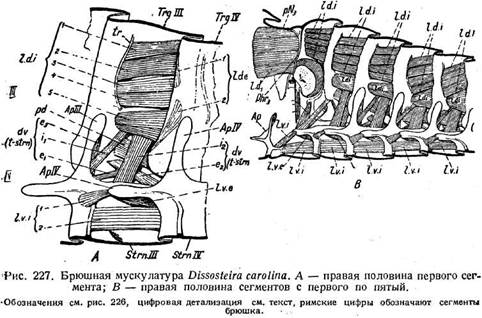
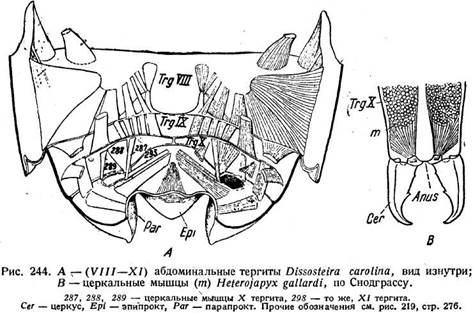
Рассмотрим мускулатуру на конкретном примере. У кобылки Dis-sosteira расположение и работа мышц довольно близко соответствуют разобранной схеме. Остановимся сначала на третьем сегменте брюшка ^рис. 227 А). Здесь имеется обширная группа парных спинных продольных мышц, внутренних и наружных. Первые (рис. 227 А, /. d. i. 1—5) -многочисленны и, как видно по их местам прикрепления (от переднего края четвертого тергита' идут ко вторичному тергальному гребешку tr на третьем тергите); они сближают сегменты друг с другом, т. е. укорачивают брюшко, и являются, следовательно, ретракторами. Самый нижний из этих мускулов является парадорсальным (pd), так как он отделен от прочих дорсо-вентральными мышцами (dv.i2). Имеется две наружных спинных мышцы (l.d.e. 1-2). Они идут почти вертикально, соединяя заднюю область третьего тергита с передним краем четвертого. Сокращаясь, они вращают •оба сегмента относительно друг к другу. Брюшная продольная группа состоит также из парных внутренних и наружных мышц. Стерниты Dis-■sostelra снабжены довольно сложными аподемами (Ар III, Ар IV), между которыми натянуты внутренние брюшные мышцы (I. и. i1,2) в числе двух.

Подобно внутренним спинным, они являются ретракторами и укорачивают брюшко при сокращении. Одна наружная брюшная мышца (l.v.e.) направляется от переднего отростка апофиза четвертого сегмента (Ар IV) назад к заднему краю третьего стер-нита. Отодвигая оба сегмента друг от друга, она растягивает брюшко и является протрактором и антагонистом внутренних брюшных мышц. Так как в спинной группе протракторов нет, то сокращение брюшных протракторов приводит к загибанию конца брюшка вверх. Дорсо-вентральная группа тоже состоит из внутренних и наружных мышц. Внутренние уже были упомянуты. В числе двух они подходят от тер-гита к стернальной аподеме (передняя —d. v. /j)h кстер-ниту (более сильная — d. v. /2). При сокращении они сближают тергит со стернитом, сжимая брюшко и способствуя выдыханию воздуха. Точно так же действуют две наружные дорсо-вентральные мышцы (du. e1} dv. e2), которые расположены очень косо, перекрещиваются друг с другом и соединяют края тергита и стернита друг с другом.. Но третья наружная мышца (dv. e.A) является антагонистом всех указанных

брюшных мышц. Она идет от верхнего конца стернальной аподемы к нижнему краю тергита. Так как первое место прикрепления расположено гораздо выше второго, то при сокращении мышц, тергит передвигается вверх, а апо-дема вместе со стернитом— вниз. Полость сегмента расширяется, что дает вдыхательный эффект в трахейной системе. Таким образом, рассмотренные мышцы обеспечивают весьма разнообразные движения, отчасти связанные с работой дыхательной системы. На рис. 227 В, где изображено пять брюшных сегментов Dissosteira, легко видеть, что расположение мускулатуры, разобранное нами для третьего сегмента, повторяется в остальных сегментах. .
Брюшная мускулатура изучена и в других группах. У поденок наблюдается максимальное сходство между личинкой и взрослым животным—особенность, характерная для примитивных групп. Наибольшие отклонения от общего плана расположения мы встречаем, во-первых, у гусениц и у личинок мух и, во-вторых, у Apterygota,
У личинок мух примитивный характер мышц сохранился только в двух брюшных тяжах (рис. 228 т. I. /.), идущих от переднегруди к заднему концу брюшка и в дорсо-вентральных мышцах. Все же остальные, которые являются производными спинных и брюшных прс-дольных мышц, приобрели диагональное расположение и образуют настоящую сетку по всему телу. Очевидно, это связано со специфическим образом жизни личинок высших мух, тогда как личиночная мускулатура низших двукрылых (Tipulidae, Psychodidae, Chironomidae, Cu-licidae, Tabanidae) весьма близка кг общему плану.
Гусеницы, со времен Лионе (1762), описавшего 1646 мышц у Cos-sus, известны сложностью брюшной мускулатуры. Снодграсс насчитывает около 150 мышц в одном брюшном сегменте гусеницы. На рис. 229 А представлены внутренние слои мышц одного сегмента Estigmene. Под ними находятся еще несколько слоев, не изображенные на рисунке. Основной план брюшной мускулатуры гусениц сильно осложнен и видоизменен. Различают двигательные мышцы и мышцы, создающие тургор. Так как почти все покровы гусеницы мягки, а в полости много крови, то тело ее можно рассматривать, как мешок, наполненный жидкостью. Давление стенок мешка на его содержимое является необходимым для сохранения формы тела и для нормальной работы мышц. Если прорезать стенку тела, то «тургоральные» мышцы сокращаются, но*так как кровь вытекает, то тургор невозможен и туловище укорачивается {1м-siocampa quercus длиною 5,7 см укорачивается до 3,8 см). Двигател1-ные мышцы при этом не сокращаются, напротив, они сгибаются и собираются в поперечные складки. Таким образом, гусеницы ушли далеко вперед по пути прогрессивной эволюции мышц.
Чрезвычайно сложна мускулатура брюшка также у некоторых Aptery-gota. Брюшные мышцы Protura сложны, но в общем укладываются в схему. Невидимому, в еще большей мере это справедливо для Campodea. Но Collembola сильно уклоняются от схемы. Наконец, у Japygidae мышцы брюшка необычайно сложны. В каждом из первых восьми сегментов брюшка насчитывается не менее сорока пар мышц (рис. 229 В), что, поэи-димому, связано с роющим образом жизни. По сложности мускулатуры Japygidae приближаются к гусеницам, но процесс усложнения шел у них совершенно иным путем. Machilidae и Lepismatidae недостаточно изучены,
§ 5. БРЮШНЫЕ ПРИДАТКИ, НЕ СВЯЗАННЫЕ С РАЗМНОЖЕНИЕМ
Брюшко имеет различные придатки, сегментарно расположенные и боль-щей частью парные. Уже это говорит в пользу их гомологизации с брюшными конечностями. Такое допущение вполне естественно еще и потому, что другие членистоногие (ракообразные, многоножки) имеют брюшные конечности, а зачатки последних нередки и в эмбриональном развитии насекомых. В дальнейшем эти зачатки либо редуцируются, либо превращаются в придатки. Однако вопрос весьма сложении лишь некоторые из брюшных придатков можно несомненно считать видоизмененными конечностями. Брюшные придатки делятся на две группы. К первой относятся придатки, служащие для движения, дыхания и других функций, не связанных с размножением. Они очень разнообразны и встречаются далеко не у всех насекомых. Вторую группу составляют половые придатки, которые имеют почти у всех насекомых общий план строения, но различны у самцов и самок.
А. Брюшные ноги Proturu
Одной из особенностей отряда Protura является присутствие рудиментарных конечностей на трех первых сегментах брюшка (рис. 217 С). У Ео-sentomon они сидят между тергитом и стернитом и состоят каждая из боль-
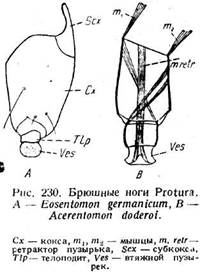
шого коксоподита и маленького телоподита (ркс. 230). Коксоподит подразделен на субкоксу, состоящую из двух маленьких пластинок (Sex), и коксу (Сх). Он приводится в движение двумя мышцами (ть т2), которые идут от тергита, одна—к переднему краю членика, другая —к заднему. Телоподит (Tip) имеет вид короткого колечка и заканчивается втяжным пузырьком (Ves), который втягивается посредством ретрак-тора (т. retr). Движения телоподита зависят от двух перекрещивающихся мышц, идущих к нему из коксоподита. Его выворачивание, очевидно, обусловлено нагнетанием крови. У Acerentomon только первая пара брюшных ног устроена таким же образом, тогда как вторая и третья утеряли расчленение и втяжной пузырек. Описан4 ные придатки несомненно соответствуют брюшным конечностям. Их существование есть доказательство родства насекомых с многоногими формами. Особенно важно, что эти конечности существуют у взрослого насекомого, а не только у личинки, как это имеет место в отношении некоторых других брюшных придатков. Следует, однако, указать, что в ноге членистоногих телоподит сильно преобладает над коксЪподитом. Здесь же телоподит развит слабо. Сейчас неясно, есть ли это редукция или, наоборот, глубокая примитивность. Вероятнее первое, но характерно, что в брюшных конечностях насекомых почти не встречаются сильно развитые телоподиты.
Б. Придатка прегенитальных сегментов Thysanura
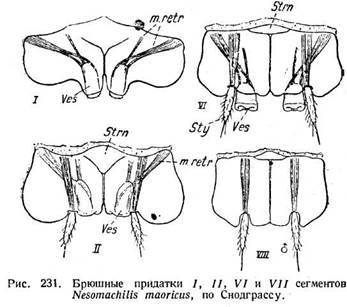
На прегенитальных сегментах Thysanura имеются характерные придатки. У Nesomachilis они состоят из коксоподита и из двух сидящих на нем
концевых частей. Коксоподит имеет форму пластинки, которая, по существу, входит в состав стенки тела; правый и левый коксоподиты сходятся по сред-
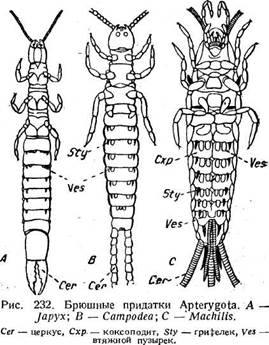
ней линии тела и в значительной мере оттесняют вперед небольшой стер-нит (рис. 231, V/). К коксоподиту идут мышцы от тергита, чем подкрепляется его гомологизация. На коксоподите сидят медиально втяжной пузырек ( Ves) и латерально-грифельный придаток, или грифелек (stylus — Sty). Пузырек приводится в действие мышцами ретракторами, которые доходят до его конца (рис. 231, II т. ret г.); мышцы грифелька доходят лишь до его основания. На разных сегментах брюшка наблюдаются различные комбинации концевых придатков. Так, на VI сегменте есть и те и другие, на / только пузырьки, на VIII у самца только грифельки. У Machilis (рис. 232 С) на четырех сегментах имеется по четыре пузырька, на двух других по ррй.УСатройеа (В) и yjapyx (А) также имеются пузырьки и грифельки. Таким образом, эти придатки в том или ином виде свойственны всем Thysanura. В отношении функции тех и других придатков есть указания, что организм
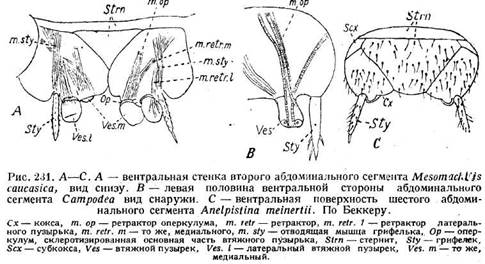
абсорбирует кислород и воду сквозь тонкие стенки пузырьков, тогда как грифельки, особенно длинные у Machilidae, поддерживают брюшко над-субстратом и, таким образом, предохраняют пузырьки от повреждений и загрязнения. Кроме того, Machilidae, как известно, будучи вообще чрезвычайно проворными и юркими насекомыми, способны к очень быстрым прыжкам. Эти последние совершаются при помощи ударов ног и хвостовых нитей и при помощи сокращений туловищной мускулатуры. Грифельки, видимо, участвуют и в этих прыжках. Природа этих придатков неясна. Казалось бы, естественно считать грифельки за телоподиты. Однако это предположение отпадает в силу того, что Machilis имеет грифельки на коксах средних и задних ног (рис. 232 С). В то же время грк-фельки свойственны не только Thysanura, но встречаются и у Pterygota> правда, лишь в числе одной пары. С другой стороны, брюшные придатки Thysanura являются исходными для понимания морфологической природы женских половых придатков высших насекомых (стр. 298). Снодграсс считает грифельки гомологами эпиподитов ракообразных. Беккер (1925),. подробно изучивший строение и мускулатуру втяжных пузырьков и гри-фельков, приходит к следующим результатам. У Campodea имеется одна пара втяжных пузырьков, .причем три ретрактора направляются от каждого пузырька вперед (рис. 231 В, т. ор), а один косо в сторону.
Такое же расположение мускулатуры наблюдается в медиальном пузырьке Machilidae (рис. 231 A, Ves. m, m.op, m.relr.m), тогда как в латеральном мускулатура иная (Ves.l, m.retr.l). Отсюда Беккер заключает, что медиальные и латеральные пузырьки имеют различное происхождение. Медиальный является гомологом телоподита абдоминальной конечности, тогда как латеральный лишь вторично вступает в связь с конечностью вследствие разрастания последней. Во втяжном пузырьке Беккер различает оперкулум (рис. 231 А, Ор), который отличается от остальной мягкой части пузырька наличием щетинок и усиленной склеротизацией. Подобно латеральному пузырьку, связь грифелька с конечностью также является вторичной и объясняется тем же выше указанным, разрастанием. Грифелек приводится в движение отводящей мышцей (рис. 231 Д, т.sty). Кроме того, склерит, который выше обозначается какс коксоподит, Беккер считает за коксу, а, например, у Anel-pisfina находит кроме коксы также и отдельную субкоксу (рис. 231 С, Сх, Sex). Эти го-мологизации, однако, не представляются вполне установленными. К вопросу о соотношении втяжных пузырьков с половыми придатками мы вернемся ниже (стр. 298, 326).
В. Брюшные придатки Collembola
Collembola имеют на брюшке три придатка: брюшную трубку, зацепку и прыгатель-ную вилку. Брюшная труб к а, или коллофор, находится на первом сегменте и представляет собою трубко-видный вырост с раздвоенным втяжным пузырьком на конце (рис. 233 А, В, Coll, Ves). Пузырек имеет два ретрактора (т. retr), отдельные для каж-дей половинки и начинающиеся от тергита. У некоторых форм (Sminthunts) каждая половинка пузырька вытягивается в свою очередь в длинную, гибкую втяжнук* трубку. Можно бы сравнить этот орган с двумя втяжными пузырьками; Protura, сросшимися по медиальной линии. Это, однако, не согласовалось бы со строением мускулатуры, ибо ретракторы пузырьков идут ке. от тергита, а от субкоксы, а тергальные мышцы управляют только» субкоксой (стр. 285). От отверстий двух пар головных желез по стер-нальной поверхности груди идет канал к брюшной трубке. Возможно,, что трубка способна прилипать к субстрату, хотя это не доказано. Беккер (1925) считает, что брюшная трубка Collembola происходит из сросшихся правой и левой конечностей первого абдоминального сегмента.
Зацепки и прыгательные вилки Collembola .Составляют один механизм. Вилка (furca —рис. 233 A, Fur) находится на четвертом сегменте и сс-стоит из рукоятки (manubrium— рис. 233 D, Man) и двух зубцов (denticuli — Dent) с маленьким придатком (Dentx) на концах. В покое она загнута вперед под брюшко и сверху не видна. Зубцами она охватывает зацепку
{tenaculum —рис. 233 С), причем зубцы вырезаны с внутренней стороны, и зацепка входит как раз в вилку (а). Зацепка представляет собою маленький раздвоенный придаток второго сегмента. Рукоятка вилки приводится в движение сгибателями и разгибателями, которые принадлежат к спинным и брюшным продольным мышцам третьего и четвертого сегментов и прикреплены к основанию вилки. Приводящие и отводящие мышцы зубцов (т. odd, m. abd) идут от стенок рукоятки соответственно
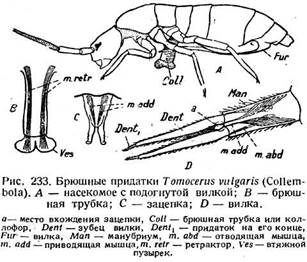
к внутреннему и наружному краям зубца. В зацепке есть только приводящие мышцы (т. add). Сокращение приводящих мышц зацепки и отводящих мышц зубцов освобождает вилку, а разгибатели вилки быстро отбрасывают ее концами назад по дуге около 180°. Насекомое получает толчок снизу и сзади и совершает длинный прыжок. Возможно, что рукоятка вилки соответствует слившимся коксам, а зубцы —разделенным телоподитам. Прыгательный аппарат характерен для отряда Collembola, но некоторые роды из семейства Poduridae лишены его.
Г. Трахейные жабры"
Каждая трахейная жабра личинки поденки состоит из двух частей. Между тергитом и стернитом находится широкая основная пластинка, подразделенная швом на две части (рис. 234 А, В, Схр). С нею подвижно сочленена собственно жабра (Вг), которая состоит из срединного стержня с двумя рядами боковых веточек, либо имеет форму округлой пластинки без разветвлений. Она приводится в движение четырьмя мышцами (рис. 234 С, тх, т2, гп3, т4), в нее входят ответвления трахейной системы. Трахейные жабры Ephemera vttlgata возникают из зачатков брюшных конечностей, имеющихся у зародыша. Поэтому основную пластинку гомологизируют коксоподиту, а жабру — грифельку Thysanura.
Личинка сетчатокрылого вислокрылки (Sialis) обладает тремя парами хорошо развитых грудных ног, за которыми следует серия придатков на первых семи сегментах брюшка (рис. 234 D). Каждый их них сидит на основной пластинке, постепенно суживается к концу, разделен на шесть члеников, подвижен и в общем имеет вид ноги. Основная пластинка, хотя и сращена с тергитом, но занимает плейральное положение, и ее полость отделена от полости тела дорсо-вентральными мышцами. От основной пластинки к основанию подвижной части идут передние и задние мышцы, но, кроме
того, первые три членика придатка имеют и собственную мускулатуру. Внутрь придатка входит трахея, которая разветвляется, начиная с четвертого членика. Основную пластинку иногда считают за коксоподит, подвижную часть за гомолог грифелька и придают особенное значение его расчленению и собственной мускулатуре, которая указывает на локомоторную функцию органа. Сходного типа трахейные жабры имеются у личинок жуков вертячек и некоторых плавунцов, но они недостаточно изучены.
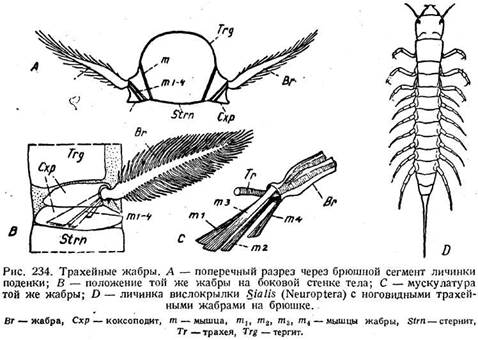
Аргументы в пользу^того, что трахейные жабры Sialis и поденок являются видоизмененными конечностями, довольно убедительны и находят поддержку в литературе. Но все же сомнение в их правильности остается. Приходится принять, что потребность в жаберном дыхании приводит то к использованию для этого конечностей, то к появлению новообразований на самых разнообразных местах тела (стр. 559—561). Конечный же результат, т. е. устройство Жабер, оказывается в обоих случаях одинаковым. Расчлененное состояние придатков у Sialis тоже говорит против сравнения с грифельками, так как последние не расчленены.
Д. Брюшные ноги гусениц и некоторые другие образования
Более убедительно сравнение с конечностями брюшных, так называемых ложных ног гусеницеобразных личинок. Очень интересный материал дают водные личинки сетчатокрылого Cbryrfa/ws,близкого к вислокрылкам. Каждый брюшной придаток у этих форм начинается с толстой выступающей сбоку основной части, которая занимает в брюшном сегменте такое же положение, как несомненная субкокса и кокса в грудном, и соответствует ко-ксоподиту (рис. 235 А, Схр). На ней сидят два придатка. В сторону торчит грифелек (Sty), напоминающий жабру Sialis, но нерасчлененный и снабженный сложной мускулатурой (рис. 235 В, т). Вниз отходит выступ, который несет пучок дыхательных нитей и три втяжные лопасти. Последние сходны с втяжными пузырьками Thysanura и приводятся в движение трех-раздельной мышцей (рис. 235 В, т. retr.), которая идет к ним от тергита.
Отличие лишь в числе пузырьков и в том, что у Thysanura мышца идет не от тергита, а от коксоподита; +1а десятом сегменте эти придатки длиннее,
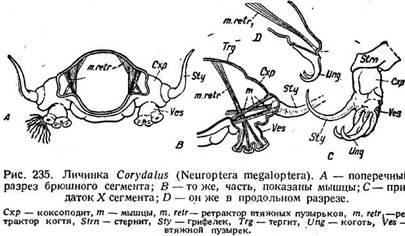
лишены дыхательных нитей, их грифелек изогнут, но зато они имеют по два больших когтя (рис. 235 D, Ung). Последние приводятся в движе-
ние сильным,ретрактором (m. retrx), идущим от тергита. Эти придатки обнаруживают близкое сходство с брюшными ногами гусениц.
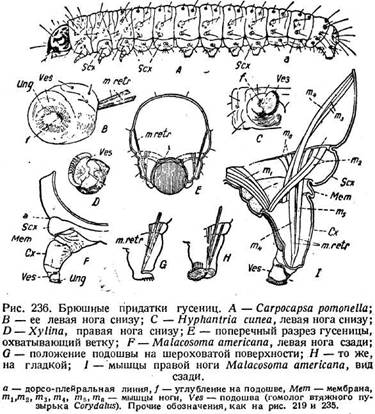
В последних можно установить присутствие субкоксы, коксы и те-лоподита. Субкокса (рис. 236 F, Sex) представляет собою часть стенки тела,

ограниченную сверху дорсо-плейральной линией (а) и имеет форму толстой лопасти или вздутия. Такие же вздутия находятся над основаниями ног в грудных сегментах. У некоторых видов расположение щетинок на субкоксах грудных и брюшных сегментов одинаково. За субкоксой следует обширная сочленовная мембрана (Мет), от которой начинается кокса (Сх). Последняя приблизительно цилиндрической формы, а наружная ее стенка нередко склеро/гизирована. Кокса и составляет собственно «ногу». Небольшая сочленовная мембрана соединяет ее с телоподитом (Ves), за которым установилось название подошвы. Он представляет собою втяжную подушку, соответствующую втяжным пузырькам Corydalus и вооруженную венчиком крючков (Ung). Нога управляется мускулатурой, делящейся на две группы. Одни мышцы приводят в движение коксу, оканчиваются на ее основном крае, а начинаются частью от средней линии брюшка (рис. 236 / тх), частью от верхнего края субкоксы (/п2), частью от боковой стенки сегмента (яг3). Общий характер их расположения подтверждает коксальную природу этой части ноги. Другая группа —это ретракторы подошвы. Из них один наиболее длинный (т4) начинается вверху около коксальной мышцы /773, два других (т5, т6) идут от верхнего края субкоксы. Все три заканчиваются на общем сухожилии, идущем у примитивных форм к центру подошвы. Длинный ретрактор напоминает Corydalus. Если оставить в стороне детали, то брюшной придаток Corydalus отличается от брюшной ноги гусеницы лишь присутствием грифелька. Даже характерные для гусениц кр^очки имеются у Corydalus, правда, лишь на последней паре придатков. "Томологизация подтверждается также эмбриологическими данными. Брюшные и грудные ноги зародыша, например Pleris rapae, развиваются совершенно одинаковым способом.
Исследование Мартыновой над гусеницей Micropteryx и личинкой панорпы указывает на существование связи между их брюшными ногами с одной стороны и рядами кожных бугорков с другой. Эти данные как будто ставят вопрос о происхождении брюшных ног в совершенно иную, чем раньше, плоскость.
Брюшных .ног у гусениц обычно бывает пять пар, четыре первые пары расположены с III по VI сегмент брюшка (т. е. с шестого по девятый сегмент туловища), и последняя пара, так называемые анальные ноги, или подталкиватели, находится jia X сегменте, так что I, II, VII, VIII и IX -сегменты брюшка лишены ног,1 В некоторых примитивных группах, например у Micropterygidae, ноги присутствуют на всех сегментах, кроме IX, и очевидно первичным состоянием является наличие ног на всех брюшных сегментах. С другой стороны, у пядениц брюшные ноги обычно сохраняются только на VI и X сегментах, что, однако, не связано с ослаблением двигательной функции. Большинство гусениц ползает, последовательно приподнимая и переставляя брюшные ноги, начиная с задней пары. Тело выгибается дугой вверх над теми ногами (рис. 237), которые в данный момент приподняты, так что по телу • проходит волна сокращения спереди назад. Когда она доходит до груди, то передвигаются и грудные ноги, после чего сзади возникает новая волна. Пяденицы передвигаются иначе. При закрепленных грудных ногах, брюшные отделяются и затем прикрепляются рядом с
1 Махотин (1940) считает подталкиватели за придатки XI сегмента и таким образом за гомологи церков.
грудными, причем тело сгибается крутой петлей вверх. Затем, при закрепленных брюшных ногах, открепляются грудные, тело распрямляется, грудные прикрепляются далеко впереди от прежнего места и таким образом один «шаг» совершен. Работа брюшных ног пядениц и их специализированной туловищной мускулатуры очень эффективна. Гусеница может, закрепившись ими, приподнять остальную часть тела и качать ее палочкообразно из стороны в сторону в поисках удобного места для прикрепления. Будучи встревожена, она замирает неподвижно в такой позе, что связано с криптизмом (стр. 456).
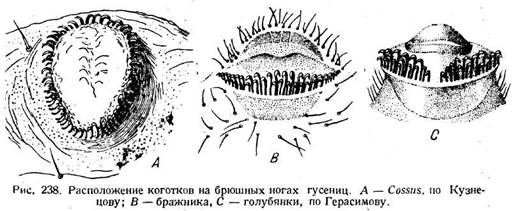
Брюшные ноги гусениц играют вспомогательную роль при приеме пищи. Обычно гусеница ест, сидя на краю листа или на стебле (стр. 60) и прочно охватывая его брюшными ногами. В связи с этим возникает вопрос о коготках брюшных ног. Основным расположением их является однорядное круговое (рис. 238 А). Круг, однако, может быть прерван либо с наружной, либо

с внутренней стороны и превращается тогда в подкову. Другие видоизменения отдельных частей круга дают еще несколько типов расположения крючков. Из них особенно распространено исчезновение латеральной половины круга. Подошва при этом утрачивает круговую формуй превращается в асимметричную лопасть с обращенными внутрь коготками на краю (рис. 236 С). Лопасть представляет собою сохранившуюся медиальную половину первичной круговой подошвы. При примитивном круговом расположении коготков их концы обращены наружу относительно центра круга, но по отношению ко всему телу они обращены наружу в латеральной половине круга и внутрь в медиальной. Поэтому при исчезновении латеральной половины сохранившиеся коготки обращены концами преимущественно к срединной плоскости тела и две ноги вместе образуют нечто вроде щипцов, вооруженных множеством коготков и прочно охватывающих стебель (рис. 236 Е), причем основное значение имеет сокращение мышцы т1. Другая серия модификаций вооружения состоит в том, что однорядное расположение крючков превращается в дву-рядное (рис. 238 В) или даже трёхрядное (рис. 238 С). Число крючков на единицу длины увеличивается вдвое или втрое, и цепкость ноги возрастает. Иногда расположение крючков прерванное (рис. 238 С). Что касается мускулатуры, то ретракторы подошвы служат для освобождения коготков от субстрата. При передвижении по шерохо-
ватой поверхности коготки поворачиваются концами вниз и зацепля* ются за субстрат (рис. 236 G), на гладкой поверхности, например на стекле, они поворачиваются концами вверх (рис. 236 Н), и тогда действие ре-тракторов, повидимому, создает вакуум, и нога присасывается. Расправление ноги происходит вследствие давления крови при расслабленном ретракторе. Известно также, что при ползании по гладким поверхностям гусеницы покрывают их тонкой паутиной, облегчая, таким образом, работу прицепного механизма.
Личинки пилильщиков гусенйцеобразны, но число брюшных ног доходит у них до восьми пар. Так, у Neodiprion (рис. 239) они находятся на 11 —VIII и X сегментах брюшка. Устройство их такое же, как у гусениц, но менее совершенно. Интересно, что у ви-дов^ живущих в растительных полостях, в гнездах или вообще
скрытно, ноги X сегмента представляют собою тонкие членистые придатки.
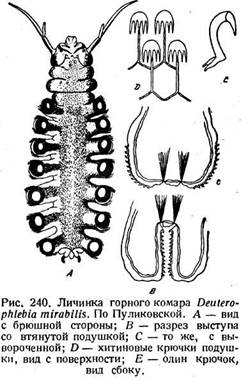
Замечательны брюшные придатки личинки горных комаров Deu-t e"r ophlebiidae, изученные
Пуликовской (1924). Личинка имеет сплюснутое тело с хорошо выраженной головой. Три грудных сегмента лишены конечностей, семь

брюшных расширены и несут каждый по паре толстых боковых выступо5 (рис. 240 А). Выступ заканчивается полушаровидной втяжной подушкой (рис. 240 В, С), которая покрыта множеством крючков, расположенных в несколько рядов. Крючки имеют сложное устройство, снабжены четырьмя остриями и обращены концами

Рис. 242. Личинка горного комара Blepharocera. А — вид с брюшной стороны; В — разрез через присоску.
7 — кряй присасывательного диска, 2—полость присоски, 3 — конусовидный поршень, Gli — первая железа, Gl% — вторая железа, т^, т% —мышцы, S. g^ —секрет второй железы.
наружу (рис 240 D, Е). Мускулатура этих придатков не изучена. Неясна также и их морфологическая природа. При их помощи личинка свободно передвигается по поверхности камней на дне горных
потоков, выдерживая сильнейший напор воды. Характерно, что при ползании личинки поворачивают все время голову вправо и влево. Возможно, что они покрывают поверхность камня каким-либо секретом, облегчающим прицепление коготков.
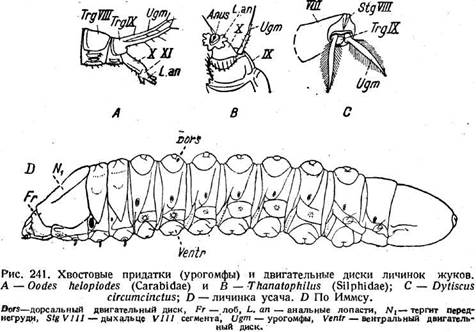
У личинок жуков также встречаются довольно разнообразные придатки двигательного характера. Для одних предлагается название урогомфов (urogomphi). Хотя они иногда сочленены (рис. 241, Ugm), но являются не конечностями, а кутикулярными отростками девятого тергита. Придатки личинок усачей имеют вид выступающих дисков на дорсальной и вентральной поверхностях брюшных сегментов (рис. 241, D, Dors, Ventr). Упираясь ими в стенки выгрызаемых в древесине галерей, личинка передвигается. Грудные ноги подвергаются при этом сильной дегенерации.
Но, быть может, наиболее замечательными брюшными придатками являются присоски личинок комаров Blepharoceridae. Эти личинки биологически сходны с личинками Deuterophlebii-dae и живут на камнях в горных потоках. Морфология их изучена недостаточно, Повидимому,
передний отдел жесткого тела личинки (рис. 242; А) соответствует первому брюшному сегменту, слившемуся с грудью и с головой. Дальше идут следующие четыре отдельных брюшных сегмента и, наконец, шестой, слитый с концевым отделом. Соответственно этому на брюшной поверхности сидят одна за другой шесть мощных присосок чрезвычайно сложного и совершенного строения (рис. 242 В). Присоска состоит из вогнутого диска, края которого загнуты вниз и внутрь и усажены крепкими щетинками (7). Середина диска образует обширное впячение (2), которое имеет сравнительно узкий вход и расширяется кверху конусообразно. Полость впячения занята почти целиком конусовидным образованием, которое можно назвать поршнем (3), ибо к нему прикреплены сильные тяжи мышц (т^, расходя-
щиеся кверху почти под прямым углом. При их сокращении поршень поднимается вверх, и в полости присоски возникает вакуум. Кроме того, имеются железы двух типов (Glu G/2) и мышцы (яь), прикрепляющиеся к периферии присоски. Действие присоски сложно. Кроме упомянутого вакуума, имеют, видимо, значение и какие-то другие факторы, так, мертвую личинку можно «присосать» к гладкой поверхности, слегка надавив ей на спину. При помощи присосок личинка превосходно удерживается на камнях под сильнейшим напором воды. Гомологизировать присоски с какими-либо другими органами не представляется пока возможным.
Е. Придатки постгенитальных сегментов
Придатки постгенитальных сегментов во многих случаях функционально объединены с таковыми прегенитальных, поэтому некоторые из них уже были описаны выше.
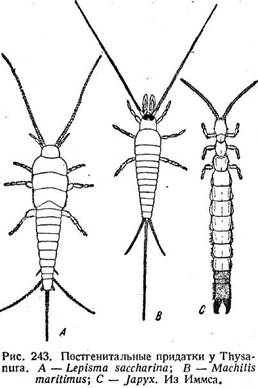
Отдельную группу составляют ц е р к и (cerci), или хвостовые нити, и хвостовой, или каудальный филамент. Они являются придатками одиннадцатого сегмента, но при его редукции могут быть вторично связаны с десятым (стр. 275). Типичные церки представляют собою длинные тонкие нити, суживающиеся к концу и состоящие из большого числа члеников. Такое строение придает им сходство с антеннами и характерно для Thysanura (рис. 243 А, В). Но по большей части они подвергаются редукции. У таракана они укорочены, хотя и сохраняют
расчленение (рис. 219 Я); у уховерток исчезает и расчленение. У большинства Holometabola церки отсутствуют, лишь самки панорпы имеют придатки, видимо, соответствующие церкам. Функционально церки являются органами чувств (стр. 653). Об их аэродинамической роли у поденок уже говорилось (стр. 247), тогда как у личинок, по Махотину (1940), они служат для плавания, чему способствует их волосяной покров, довольно обильный у некоторых видов. О их функции у Machilidae сказано выше (стр. 286). У Japygidae и у уховерток они превращаются в хватательные щипцы (рис. 243 С, 244 В). Мускулатура церков у Dissosteira состоит из четырех пар дорсальных мышц, причем три (рис. 244 А, 287, 288, 289) начинаются от X и лишь одна (293) от XI сегмента. Однако мышечная связь церков с X сегментом является, видимо, вторичной и не считается доводом против их принадлежности к XI сегменту. В эмбриогенезе ряда прямокрылых церки возникают на XI сегменте в виде бугорков, которые ничем не отличаются от зачатков конечностей предыдущих сегментов. Этим окончательно устанавливается их природа.
Каудальный филамент сходен по строению с церками. Он может достигать приблизительно такой же длины, как и церки, и тогда насекомое имеет три приблизительно одинаковых нити. Но нередко при хорошо развитых церках филамент может быть укорочен (рис. 219 С) или отсутствует (рис. 219 D).
Дата добавления: 2019-09-30; просмотров: 623;
Поиск по сайту
Узнать еще
- Andantino con moto А. Бородин. Для берегов отчизны дальней
- B) в угле Интинского месторождения и продуктах его сжигания.
- F. Построение диаграмм сумм рангов
- I. Государственный бюджет и его структура. три состояния государственного бюджета.
- I. Общая характеристика категории состояния как часть речи
- I. Товар и его свойства.
- II. ГОСУДАРСТВЕННЫЙ ДОЛГ И ПУТИ ЕГО ПОГАШЕНИЯ
- II. Движение поездов на однопутных перегонах

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
