Освобождение медиаторов
В пресинаптическом нервном окончании в синаптических везикулах аккумулируются медиаторы (трансмиттеры). Потенциал действия, достигая нервного окончания, деполяризует его мембрану. Деполяризация вызывает открытие кальциевых каналов, по которым в окончание входит ток ионов Са2+. Ионы Са2+ вызывают процесс слияния везикул с пресинаптической мембраной с последующим открытием их и выбрасыванием своего содержимого в синаптическую щель.
Для более полного описания процесса синаптической передачи рассмотрим холинергический синапс, в котором в качестве медиатора освобождается ацетилхолин (Ach) (рис. 1.5, 1.6).
Синаптические везикулы фиксируются большей частью посредством протеина синапсина, который локализован на цитоплазматической поверхности каждой везикулы, к протеину спектрину, расположенному на волокнах F-актина цитоскелета, образуя тем самым трансмиттерный резервуар. Меньшая часть везикул связана специфическими протеинами с внутренней стороной пресинаптической мебраны. Это взаимодействие осуществляется посредством белка мембраны везикулы синаптобревина и белка пресинаптической мембраны синтаксина. Именно эти везикулы непосредственно поставляют медиатор для очередного выброса.
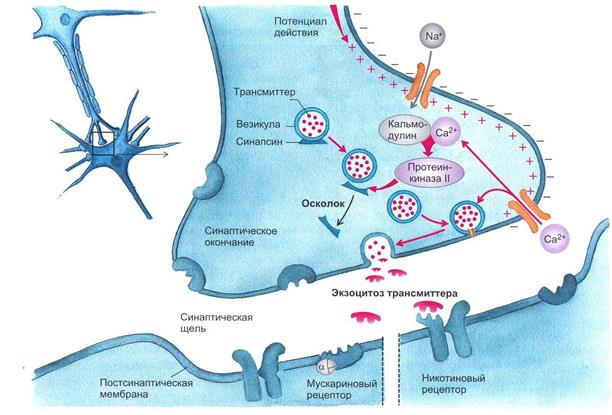
Рис. 1.5. Механизм освобождения трансмиттера из пресинаптического окончания.
Потенциал действия, пришедший по аксону в пресинаптическую область, деполяризует мембрану. Открываются потенциалуправляемые Са+-каналы. Повышенная концентрация интратерминального Са+ открывает везикулы, лежащие на пресинаптической мембране клетки, что приводит к экзоцитозу их содержимого в синаптическую щель
(рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004)
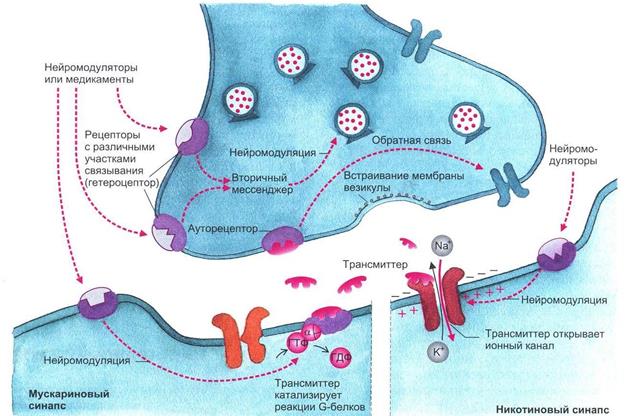
Рис. 1.6. Взаимодействие трансмиттерных молекул с их специфическими рецепторами.
В правой части рисунка представлена постсинаптическая мембрана, имеющая никотиновый рецептор, ионный канал которого открывается сам при помощи лиганда; в левой части показана постсинаптическая мембрана, обладающая мускариновым рецептором. В этом случае ионный канал открывается только при помощи каскада химических реакций (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред.
А. Камкина и А. Каменского. М., 2004)
Если потенциал действия достиг пресинаптической области и в пресинаптическом окончании концентрация Са2+ поднялась до необходимого уровня, то происходят следующие процессы. Са+2 связывается с протеином, входящим в состав мембран везикулов, – синаптотагмином. Это приводит к тому, что мембрана везикулы раскрывается. Одновременно комплекс полипептида синаптофизина сливается с неиндефицированными протеинами пресинаптической мембраны. При этом возникает пора, через которую осуществляется регулируемый экзоцитоз, т.е. выход трансмиттера в синаптическую щель. Протеин везикулы rab3A регулирует этот процесс. В одной везикуле сосредоточено примерно 6000-8000 молекул трансмиттера. Это наименьшее количество трансмиттера, освобожденного в синаптическую щель, называется «один квант трансмиттера». В совокупности локальная концентрация медиатора в синаптической щели после его освобождения относительно высока и находится в миллимолярном диапазоне.
Повышенный уровень ионов Ca2+ в пресинаптическом окончании активирует Са2+-кальмодулинзависимую протеинкиназу П. В пресинаптическом окончании этот фермент фосфорилирует синапсин. После этого нагруженные трансмиттером везикулы освобождаются от цитоскелета и перемещаются на пресинаптическую мембрану для осуществления дальнейшего цикла, описанного выше. Главную роль для процесса выбрасывания медиатора в синаптическую щель играет входящий ток Са2+. Деполяризация окончания лишь открывает кальциевые каналы. Ионы Са2+ служат здесь в качестве вещества-посредника (вторичного мессенджера), которое запускает механизм слияния везикул с пресинаптической мембраной.
Повышение концентрации экстрацеллюлярного Са2+ повышает входящий ток ионов Са2+, что увеличивает освобождение медиатора. Искусственное повышение концентрации экстрацеллюлярного Mg2+ посредством замещения им ионов Са2+ ведет к снижению входящего тока Са2+ и тем самым уменьшению освобождения трансмиттера. G-белки, вероятно, также участвуют в управлении везикулярным экзоцитозом. После прекращения пресинаптического потенциала действия ионы Са2+ удаляются из пресинаптической области посредством активного ионного транспорта с участием Са2+-АТФазы и 3Na+/Ca2+-обменника.
Многоразовое сильное возбуждение пресинаптического нейрона ведет к увеличению концентрации ионов Са2+ и тем самым к повышению освобождения трансмиттера. Этот процесс называется синаптическим или посттетаническим потенцированием (после тетанического раздражения). Предполагают, что механизм посттетанического потенцирования лежит в основе памяти. Подобный процесс широко представлен в гиппокампе. Механизмы синаптического потенцирования повышают эффективность синапса. Входящим током ионов Са2+ можно управлять, влияя на Са2+-каналы. Они могут быть более или менее частыми, закрытыми или открытыми. Возможно фармакологическое воздействие на каналы. Например, конотоксин – яд улитки – блокирует каналы Са2+. Кроме того, ионы Mg2+, вытесняя ионы Са2+, уменьшают освобождение медиатора. Опорожнение везикул также может блокироваться посредством ядов. Так, ботулинический токсин при мясном отравлении действует на пресинаптическую мембрану, препятствуя скоплению везикул на ней за счет уменьшения необходимого для этого процесса протеина. Подобным образом действует тетанустоксин, который в переднем роге спинного мозга прекращает торможение мотонейрона клетки Реншоу, что приводит к судорогам.
Дата добавления: 2016-07-27; просмотров: 5304;
Поиск по сайту
Узнать еще
- А. Освобождение дыхательных путей
- Борьба польского народа за национальное освобождение в XIX в.
- Великое Московское княжество и освобождение русских земель от монгольского ига. Формирование центральных органов власти.
- Механизм действия медиаторов
- Освобождение и отстранение опекунов и попечителей от исполнения ими своих обязанностей. Прекращение опеки и попечительства
- Освобождение от административной ответственности
- Освобождение от действия электрического тока

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
