Структуры прокариотной клетки
Клетка прокариот имеет все структурные компоненты, присущие любой клетке. Среди структур бактериальной клетки различают:
основные структуры – клеточную стенку, цитоплазматическую мембрану, цитоплазму с различными цитоплазматическими включениями и нуклеоид;
временные структуры – капсулу, жгутики, фимбрии, у некоторых представителей бактерий – эндоспоры.
Клеточная стенка. Клеточная стенка является обязательным структурным элементом бактериальной клетки, исключение составляют микоплазмы и L-формы бактерий. Обладая высокой степенью эластичности и упругости, она выдерживает внутриклеточное давление. Клеточная стенка служит механическим барьером между протоплазмой и внешней средой, придает клеткам определенную форму, определяет способность удерживания или вымывания красителей, дает возможность клетке существовать в гипотонических растворах.
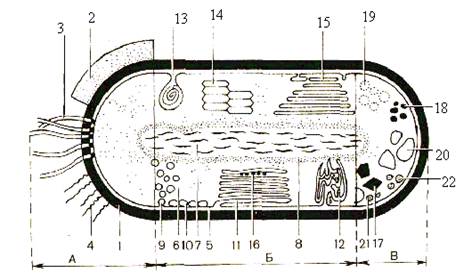
Рис. 1. Схематическое строение прокариотной клетки. А – поверхностные структуры: 1 – клеточная стенка; 2 – капсула, или слизистый слой;
3 – жгутики; 4 – фимбрии; Б – цитоплазматические структуры: 5 – цитоплазматическая мембрана; 6 – цитоплазма; 7 – нуклеоид; 8 – рибосомы;
9 –хроматофоры; 10 – везикулы; 11 – пластинчатые тилакоиды; 12 – трубчатые тилакоиды; 13 – мезосома; 14 – аэросомы; 15 – ламеллярные структуры; 16 – карбоксисомы; В – запасные вещества: 17 – полифосфаты;
18 – полисахариды; 19 – поли-b-оксимасляная кислота; 20 – включения серы; 21, 22 – жировые капли (Г. Шлегель, 1972)
Поверхностные макромолекулы внешней стороны клеточной стенки выполняют разнообразные функции, такие, как:
– специфическая рецепторная для фагов и колицинов;
– антигенная роль;
– функция межклеточного взаимодействия при коньюгации и при взаимодействии с клетками тканей высших организмов.
Клеточная стенка грамотрицательных (грациликутных) значительно расширяет круг функций, так как осуществляет роль дополнительного клеточного барьера, имеет дополнительные специфические и неспецифические каналы (диффузные поры), препятствует проникновению в клетку токсических веществ.
Без специальных методов окраски рассмотреть клеточную стенку бактерий под световым микроскопом не удается. На долю клеточной стенки приходится от 5 до 50% сухой массы клетки. Толщина данной структуры составляет 10–80 нм. Клеточной стенкой или клеточной оболочкой называют структуры, расположенные над цитоплазматической мембраной. Различают также поверхностные дополнительные структуры (капсулы, жгутики, фимбрии и т.п.).
По химическому составу клеточная стенка прокариот коренным образом отличается от оболочки клеток эукариот. Основным компонентом клеточной стенки бактерий является муреин, относящийся к классу пептидогликанов. Муреин – гетерополимер, образованный чередующимися остатками N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенными b-1,4-гликозидными связями. N-ацетилмурамовая кислота соединена с пептидом, в состав которого входят 4–6 различных аминокислот. Основу пептидного компонента муреина составляют тетрапептиды, образованные обычно L-аланином, D-глутаминовой кислотой, специфической для прокариот мезодиаминопимелиновой кислотой (мезо- ДАП) и D-аланином. У некоторых бактерий мезодиаминопимелиновая кислота замещена на L-лизин, либо L- или D-орнитин, либо на 2,4-диаминомасляную кислоту.
По содержанию муреина и специфике дополнительных компонентов, включенных в муреиновую сеть, все бактерии подразделяются на две группы: грамположительные (фирмакутные) и грамотрицательные (грациликутные).
В 1884 г. X. Грам предложил метод окраски бактерий, вошедший в практику микробиологии как один из диагностических признаков. Метод основан на различной способности микробов удерживать красители трифенилметанового ряда – кристаллвиолет и генцианвиолет в клетке. При окрашивании фиксированных мазков раствором фиолетового красителя и закреплении его раствором йода в калий-йод, грамположительные бактерии образуют стойкое соединение красителя с йодом и при последующей обработке мазков спиртом или ацетоном не раскрашиваются, сохраняя фиолетовую окраску. Грамотрицательные бактерии не образуют стойкого соединения красителя с йодом, полностью обесцвечиваются спиртом или ацетоном, и мазки их подлежат дополнительной окраске, обычно водным раствором фуксина. Способность микроорганизмов окрашиваться по методу Грама или утрачивать окраску объясняется спецификой химического состава и ультраструктуры их клеточной стенки.
Клеточная стенка грамположительных бактерий достаточно массивна, толщина ее достигает 20–80 нм. Она имеет гомогенную губчатую структуру, пронизанную порами, и плотно прилегает к цитоплазматической мембране. Муреин в клеточной стенке грамположительных бактерий составляет от 50 до 90% ее сухой массы. С муреином связаны тейхоевые кислоты. Они представляют собой полимеры трехатомного спирта глицерина или пятиатомного спирта рибита, остатки которых соединены фосфодиэфирными связями. Одна молекула тейхоевой кислоты обычно включает от 7 до 15 спиртовых остатков. Свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилглюкозамина или N-ацетил-галактозамина. Глицеринтейхоевые кислоты нередко связаны с липидом, находящимся в цитоплазматической мембране. Тейхоевые кислоты оказывают влияние на катионный обмен клетки. У некоторых бактерий они принимают участие в регуляции активности автолитических ферментов – гидролаз, способных разрушать собственную оболочку. В составе клеточной стенки грамположительных бактерий в небольшом количестве обнаружены полисахариды, белки и липиды. Оболочка фирмакутных – эффективный ионообменник с высокой поглотительной способностью и может содержать большое количество катионов. Поглотительная способность оболочек фирмакутных сопоставима с лучшими промышленными катионитами. У большинства фирмакутных в составе оболочек присутствуют дополнительные структурные компоненты, образованные полисахаридами, белками или гликопротеидами. Нередко одиночно расположенные молекулы белков образуют дополнительную структуру на поверхности основного слоя клеточной стенки. Например, молекулы белка А стафилококков формируют одномолекулярную антипараллельную структуру, обладающую высокой биологической активностью. Белок А вызывает агглютинацию эритроцитов, угнетает фагоцитоз, комплемент, препятствует трансформации лимфоцитов, является иммунодепрессантом. У стрептококков группы А белок М образует ворсинки из спаренных молекул, связанных с муреином оболочки. Белок М обеспечивает связь в цепочке кокков, а также взаимодействие с клетками эпителия. По своему строению белок М очень близок к мышечному белку позвоночных тропомиозину, поэтому организм не вырабатывает против него антитела. Белок М является иммунологическим мимикрантом.
В наиболее сложно устроенных оболочках фирмакутных на срезах выявляются два электронно плотных слоя, разделенных слоем с меньшей электронной плотностью для электронов, пронизанных фибриллярными мостиками, имеющие на поверхности структуры белков.
Клеточная стенка грамотрицательных (грациликутных) бактерий многослойна, отличается структурной и функциональной сложностью, толщина ее составляет 14–17 нм. По химическому составу она более разнообразна. Эта группа прокариот отражает один из прогрессивных путей эволюции бактерий. Для них характерно наличие пространства между ЦПМ и клеточной стенкой, появление дополнительной внешней мембраны с большей структурной сложностью, меньшая доля муреина, входящего в состав муреинового мешка – саккулы.
Внешний слой клеточной стенки – наружная мембрана образована фосфолипидами, липополисахаридами – ЛПС, липопротеидами и белками. Основной фракцией наружной мембраны являются липиды, составляющие в среднем 22% сухой массы клеточной стенки.
По строению наружная мембрана имеет типичную трехслойную организацию, характерную для элементарных мембран. Наружная мембрана отличается большей жесткостью и более сложным химическим составом. Основу внешней мембраны составляют липиды (ПЛС, липопротеины, фосфолипиды) и белки. ПЛС составляет около 30–40% поверхности мембраны, локализован во внешнем лепестке мембраны. ЛПС – вещество с высокой антигенной активностью, О-антиген или соматический бактериальный антиген, вызывает выработку антител иммунной системой макроорганизма. ЛПС сальмонелл состоит из липида А и гетерополисахаридной структуры, в которой выделяют О-антигенные цепи и ядро. О-антигенные или О-специфические цепи состоят из повторов олигосахаридов или гетерополисахаридов уникальной структуры. Эти цепи выступают над клеточной стенкой, образуя ворсинки рецепторного назначения, активно взаимодействующие с окружающим миром. О-антигенные цепи клубеньковых бактерий находят корни бобовых симбионтов.
О-антигенные цепи определяют устойчивость бактерий к действию фагоцитов, они часто мутируют и таким образом приобретают устойчивость к действию специфических антител, что дает преимущества патогенным бактериям.
Ядро структуры ЛПС является более стабильной структурой. Иногда выделяют внешнюю и внутреннюю части. Внешняя часть состоит из гексоз разного типа (Д-глюкозы. Д-галактозы, N-ацетилглюкозоамин). Внутренняя часть состоит из гептоз, характерных ЛПС – 2-кето-3дезокси-Д-манно-октоната (КДО) и других гептоз. Формирование полной структуры ЛПС у бактериального организма определяет некоторые культуральные свойства. Бактерии, имеющие полную структуру ЛПС образуют колонии с блестящей поверхностью, а с нарушением в структуре ЛПС – колонии с матовой поверхностью, обладают рядом дефектов внешней мембраны, часто отличаются селективными преимуществами по сравнению с нормой по ЛПС.
Липид А является мембранным носителем для ЛПС и представляет собой фосфолипид. Центральную часть липида А составляет дисахарид, состоящий из двух фосфорилированных Д-глюкозоаминовых остатков. Глюкозоамины ацилированы жирнокислотными остатками.
ЛПС большинства бактерий токсичен для животного организма, то есть является эндотоксином. ЛПС вызывает лихорадку, лейкоцитоз, токсический шок и даже смерть.
Липопротеин Брауна – основной компонент внешней мембраны, имеет молекулярную массу около 7000, состоит из 58 аминокислот.
В липопротеине повторяется 15 аминокислот. Первичная структура расшифрована Брауном, предполагается эволюционное закрепление дупликации гена. Отсутствие или нарушения в строении липопротеина влияют на функции внешней мембраны.
Белки основы внешней мембраны или порины составляют до 80% белков мембраны, выполняют функции формирования гидрофильных пор. Известно более 10 видов белков основы, но одновременно в клеточной оболочке бывает 2–3, но не более 5 видов. Молекулы поринов формируют тримеры с образованием гексагональной сетки матрикса мембраны. Предположительно три молекулы порина образуют водную пору диаметром 2 нм. Каждая молекула порина стабилизирована тремя молекулами липопротеина, одна из каторых ковалентно соединена с муренином. Белковый матрикс оболочек грациликутных определяет прочность и форму клеток. Белки основы выполняют важную роль в транспорте элементов питания, служат рецепторами для фагов и колицинов. Белки порины играют большее значение для прокариот естесственных мест обитания, чем для культивируемых.
Вторую группу белков составляют минорные белки внешней мембраны. Они выполняют транспортные и рецепторные функции. Полифункциональность – основное характерное свойство белков мембраны. Белки определяют культуральные свойства бактерий.
Внешняя мембрана содержит двухвалентные катионы, которые удаляют с помощью ЭДТА или воды. Внешняя мембрана обладает трансмембранным потенциалом, она отрицательно заряжена со стороны периплазмы. Величина потенциала зависит от катионного состава среды; обычно оценивают в 20–30 Мв.
Периплазматическое пространство составляет около 10 нм, объем зависит от условий среды. Периплазма содержит муреиновый слой (саккулу) и раствор белков и олигосахаридов. Толщина муреинового слоя
1,6–3 нм, что соответствует одному-трем слоям муреина. Белки периплазмы двух типов. Встречаются гидролитические ферменты и транспортные белки. Возможно, что транспортные белки периплазмы ассоциированы с белками ЦПМ, которые обуславливают транспорт субстратов. Олигосахариды играют решающую роль в осморегуляции клетки.
У некоторых видов грациликутных на внешней мембране имеются белковые молекулы, образующие сетчатую структуру, несущие дополнительные зашитные функции.
Дата добавления: 2016-07-27; просмотров: 4951;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. Поверхностный аппарат клетки.
- ID_структуры . ID_поля
- А) Определение резистентности грудной клетки
- Алгоритм расчета структуры изображения с использованием функции размытия линии.
- Альтернативные структуры ДНК
- Анаболическая система клетки
- Анализ ассортимента и структуры продукции

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
