Наследственность, сцепленная с полом
Половые различия раздельнополых организмов связаны с различиями между их хромосомами, однако они по-разному проявляются у организмов разных видов.
У насекомых рода Protenor различия между хромосомами мужских и женских особей заключаются в том, что у мужских особей хромосомы представлены нечетным (меньшим) количеством (13), тогда как у женских особей — четным (14), т. е. количеством, большим на одну хромосому, которая является добавочной. Эта добавочная хромосома и определяет пол унаследовавшей ее особи в результате оплодотворения яйцеклетки мужской половой клеткой.
Однако у многих раздельнополых растений, беспозвоночных, животных, рыб, птиц и всех млекопитающих различия между хромосомами мужских и женских особей связаны не с количеством хромосом, а с их качественным составом. Например, соматические клетки мужских и женских особей D. melanogaster несут по четыре пары хромосом. Как у самцов, так и самок три пары хромосом являются одинаковыми и их называют аутосомами, но члены четвертой пары у самцов и самок неодинаковы по строению, ибо у самок оба члена этой пары являются прямыми палочковидными образованиями (Х-хромосомы), тогда как у самцов один член пары является прямым палочковидным образованием (Х-хромосома), а второй — изогнутым (Y-хромосома). Все одинаковые хромосомы у самцов и самок называют аутосомами (А), тогда как хромосомы Х и Y получили название половых хромосом. Все яйцеклетки плодовой мушки несут четыре хромосомы (ЗА+Х), из которых три являются аутосомами, четвертая — Х-хромосомой. Напротив, сперматозоиды также обладают четырьмя хромосомами, но они наполовину несут Х-хромосому (ЗА+Х) и наполовину — Y-хромосому (3A+Y). Оплодотворение любой яйцеклетки сперматозоидом, обладающим Х-хромосомой, дает начало зиготе женского типа (6А+ХХ), тогда как оплодотворение любой яйцеклетки сперматозоидом, обладающим Y-хромосомой, дает начало зиготе мужского типа (6A+XY).
У птиц самцы в соматических клетках содержат хромосомы XX, самки — хромосомы XY. Детерминирование пола у птиц сходно с этим явлением у D. melanogaster.
У человека хромосомы в соматических клетках женщин представлены 22 парами аутосом и парой половых хромосом XX (22АА+ХХ), тогда как в соматических клетках мужчин — 22 парами аутосом и парой половых хромосом XY (22AA+XY). Следовательно, каждая яйцеклетка несет 22 аутосомы и половую хромосому X, но сперматозоиды наполовину являются клетками, несущими 22 аутосомы и одну хромосому X, и наполовину клетками, несущими 22 аутосомы и одну хромосому Y. На хромосоме Y локализован ген длиной в 230 000 пар азотистых оснований, детерминирующий развитие тестисов (ZFY-секс-детерминирующий район Y). С геном ZFY связан также Y-хромосомный ген SRY, который тоже вовлечен в контроль развития тестисов и, кроме того, в контроль формирования костей. Кроме того в дополнение к гену существуют несколько аутосомных генов и один Х-сцепленный генный локус. Эти гены связаны с дефектами в развитии тестисов.
У человека и млекопитающих пол потомства также зависит от сперматозоида, оплодотворяющего яйцеклетку. Проникновение в яйцеклетку ядерного содержимого сперматозоида, несущего Х-хромосо-му, даст зиготу женского типа. В соматических клетках женских индивидов синтез мРНК идет лишь на одной хромосоме Х (в одних клетках на материнской хромосоме X, в других — на отцовской). Проникновение в яйцеклетку сперматозоида с Y-хромосомой даст зиготу мужского типа. Мужские индивиды являются гетерозиготны-ми (гетерогаметными) по полу, тогда как женские — гомозиготными (гомогаметными). Таким образом, пол генетически контролируется.
Однако у животных ряда видов пресмыкающихся пол контролируется не генотипом, а средой. Например, у многих черепах пол детерминируется температурой инкубации отложенных яиц (женские особи развиваются при повышенных температурах инкубации, мужские — при пониженных). А у аллигаторов, крокодилов и ящериц отдельных видов мужской пол детерминируется повышенными температурами. Детерминирование пола средой у позвоночных является исключением из общего правила генетического контроля пола.
Еще в 1910 г. Т. Морган (1886-1945) установил, что половые хромосомы участвуют также в контроле и ряда признаков. В его экспериментах, заключающихся в скрещиваниях белоглазых дрозофил-самок с красноглазыми дрозофилами-самцами, оказалось, что гибридное потомство состояло из красноглазых самок и белоглазых самцов. Однако среди гибридов встречались отдельные красноглазые самцы и белоглазые самки, появление которых было неожиданным, исходя из особенностей передачи хромосомы Х и нахождения на ней локуса гена, контролирующего белый цвет глаз. Анализируя этот случай, К. Бриджис (1916) предположил, что «необычные» белоглазые самцы несут по две Х-хромосомы, унаследовав их от своих «матерей», тогда как «необычные» красноглазые самцы имеют одну Х-хромосому, унаследовав ее от «отцов». Объясняя это предположение, К. Бриджис допустил, что исходные белоглазые самки продуцировали «необычные» гаметы, содержащие либо две хромосомы XX, либо ни одной хромосомы Х (наряду с гаметами, обычно содержащими по одной хромосоме X) в результате нерасхождения при мейозе их хромосом XX. Если такие «необычные» гаметы подвергались оплодотворению сперматозоидами красноглазых самцов, то это приводило к образованию зигот четырех типов: а) белоглазых самок, б) красноглазых самцов, в) особей с 3 хромосомами Х нежизнеспособных, г) особей без хромосомы Х (также нежизнеспособных). Назвав это явление первичным нерасхождение хромомсом (рис. 128). К. Бриджис затем цитологически показал, что в соматических клетках «необычных» белоглазых самок F, вместо хромосом XX присутствуют две Х и одна Y-хромосома (XXY).
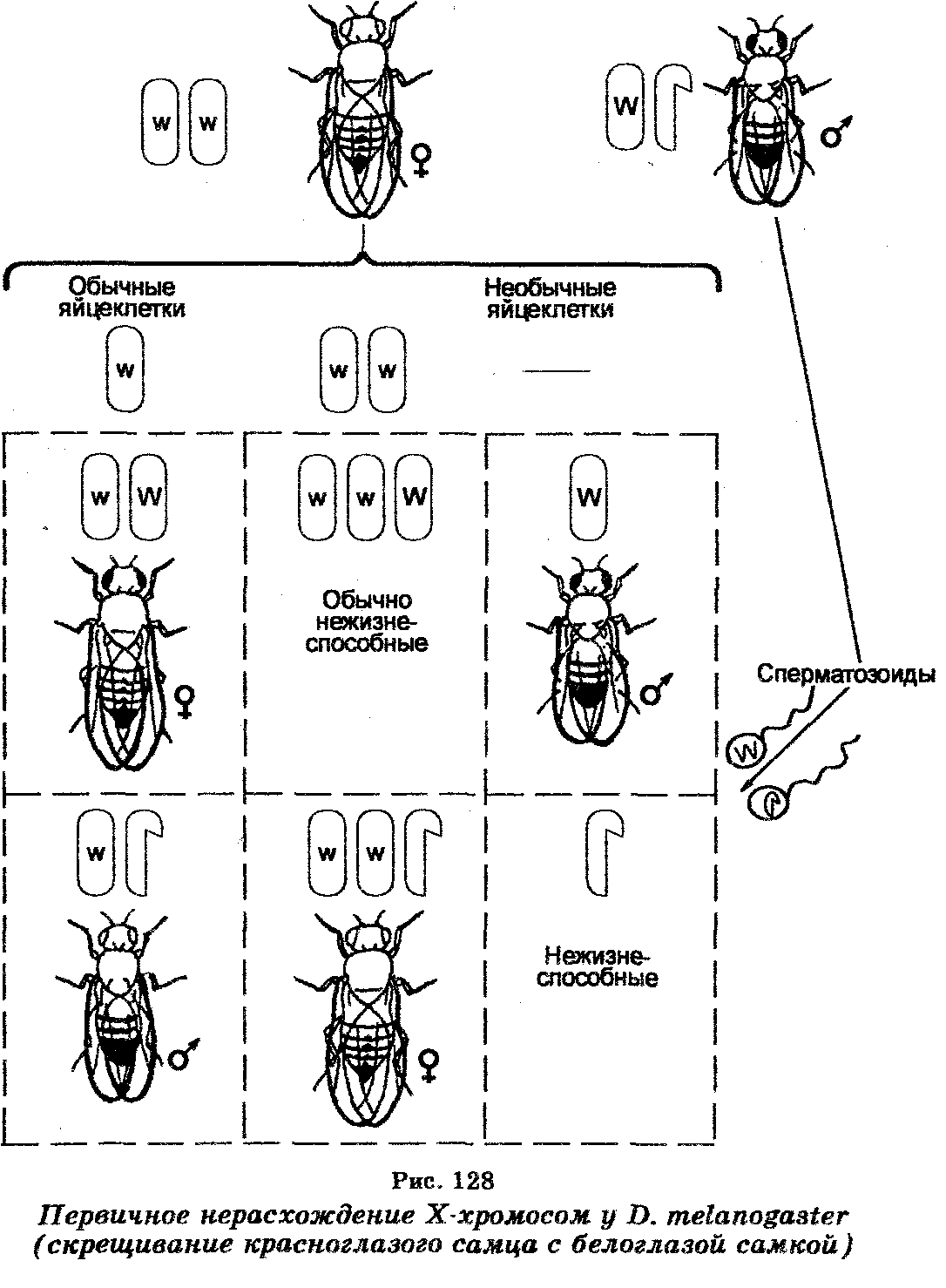
«Необычные» красноглазые самцы, не имеющие хромосомы Y, т. е. моносомики ХО, оказались в дальнейших экспериментах стерильными, тогда как белоглазые самки XXY были плодовитыми. Скрещивая последних с нормальными красноглазыми самцами и анализируя их потомство F1, К. Бриджис открыл вторичное нерасхождение хромосом при мейозе у самок XXY (рис. 129), поскольку среди организмов F, 96% самок имели красные глаза и 4% — белые, тогда как среди самцов 96% были белоглазыми и 4% — красноглазыми. В этом случае при мейозе самки XXY продуцируют гаметы четырех типов, в которых хромосомы Х и Y распределяются неодинаково (яйцеклетки с хромосомой X, яйцеклетки, имеющие по одной хромосоме Х и Y, яйцеклетка с двумя хромосомами Х и яйцеклетки лишь с хромосомами Y).
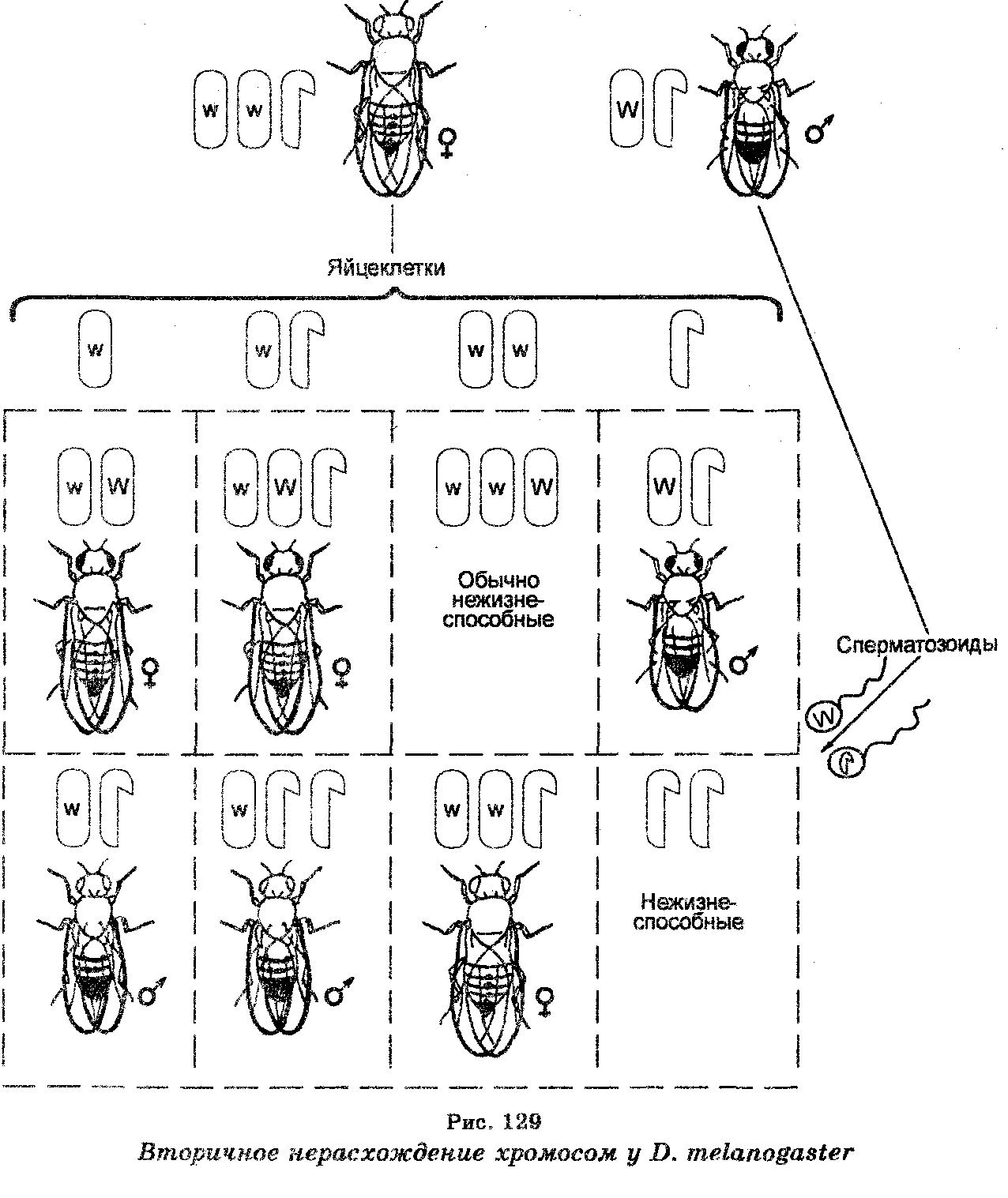
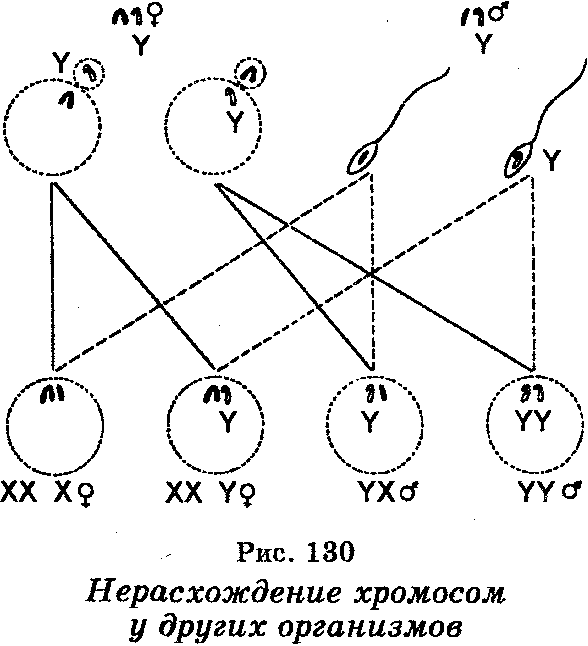
Оплодотворение яйцеклеток каждого из этих типов сперматозоидами нормального красноглазого самца приводило к образованию зигот восьми типов. Однако эти зиготы будут формироваться не с одинаковой частотой, поскольку мейоз у самок XXY связан с тем, что расхождение двух хромосом Х имеет место лишь в 92% овоцитов Объясняя вторичное нерасхождение хромосом, К. Бриджис допустил, что все белоглазые и отдельные красноглазые самки должны обладать не только парой хромосом X, но и хромосомой Y, тогда как красноглазые самцы в отличие от таких самцов, отмечаемых при первичном нерасхождении, должныиметь Y-хромосому и быть плодовитыми. Что касается отдельных белоглазых самцов, то они должны обладать одной хромосомой Х и двумя хромосомами Y. Эти допущения К. Бриджис доказал цитологически, найдя в соматических клетках дрозофилы хромосомы во всех сочетаниях, приведенных выше. Это открытие было показано на примере и других организмов (рис. 130).
 Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII).
Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII).
Дата добавления: 2016-05-30; просмотров: 1859;
Поиск по сайту
Узнать еще
- Групповое консультирование несовершеннолетних по вопросам общения с другим полом
- Имеющих трудности в общении с другим полом
- Курсивом выделены патологические признаки наследования, сцепленные с полом
- Наследование признаков сцепленных с полом.
- Наследование признаков, сцепленных с полом
- Наследование признаков, сцепленных с полом.
- НАСЛЕДОВАНИЕ, СЦЕПЛЕННОЕ С ПОЛОМ
- Наследственность, изменчивость и среда

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
