The Adaptive Immune System Requires Cooperation Between Lymphocytes and Antigen-Presenting Cells
Adaptive Immunity and Cells of Immune System
Adaptive immunity is capable of recognizing and selectivelyeliminating specific foreign microorganisms and molecules(i.e., foreign antigens). Unlike innate immune responses, adaptive immune responses are not the same in all membersof a species but are reactions to specific antigenic challenges.
Adaptive immunity displays four characteristic attributes:
· Antigenic specificity
· Diversity
· Immunologic memory
· Self/nonself recognition
The antigenic specificityof the immune system permits it to distinguish subtle differences among antigens. Antibodies can distinguish between two protein molecules that differ in only a single amino acid. The immune system is capable of generating tremendous diversity in its recognition molecules, allowing it to recognize billions of unique structures on foreign antigens. Once the immune system has recognized and responded to an antigen, it exhibits immunologic memory; that is, a second encounter with the same antigen induces a heightened state of immune reactivity. Because of this attribute, the immune system can confer life-long immunity to many infectious agents after an initial encounter. Finally, the immune system normally responds only to foreign antigens, indicating that it is capable of self/nonself recognition. The ability of the immune system to distinguish self from nonself and respond only to nonself molecules is essential, for, as described below, the outcome of an inappropriate response to self molecules can be fatal.
Adaptive immunity is not independent of innate immunity. The phagocytic cells crucial to nonspecific immune responses are intimately involved in activating the specific immune response. Conversely, various soluble factors produced by a specific immune response have been shown to augment the activity of these phagocytic cells. As an inflammatory response develops, for example, soluble mediators are produced that attract cells of the immune system. The immune response will, in turn, serve to regulate the intensity of the inflammatory response. Through the carefully regulated interplay of adaptive and innate immunity, the two systems work together to eliminate a foreign invader.
The Adaptive Immune System Requires Cooperation Between Lymphocytes and Antigen-Presenting Cells
An effective immune response involves two major groups of cells: T lymphocytes and antigen-presenting cells. Lymphocytes are one of many types of white blood cells produced in the bone marrow by the process of hematopoiesis. Lymphocytes leave the bone marrow, circulate in the blood and lymphatic systems, and reside in various lymphoid organs. Because they produce and display antigenbinding cell-surface receptors, lymphocytes mediate the defining immunologic attributes of specificity, diversity, memory, and self/nonself recognition. The two major populations of lymphocytes – B lymphocytes (B cells)and T lymphocytes (T cells) – are described briefly here and in greater detail in later chapters.
B LYMPHOCYTES
B lymphocytes mature within the bone marrow; when they leave it, each expresses a unique antigen-binding receptor on its membrane (Figure 1 a). This antigen-binding or B-cell receptor is a membrane-bound antibody molecule.Antibodies are glycoproteins that consist of two identical heavy polypeptide chains and two identical light polypeptide chains. Each heavy chain is joined with a light chain by disulfide bonds, and additional disulfide bonds hold the two pairs together. The amino-terminal ends of the pairs of heavy and light chains form a cleft within which antigen binds. When a naive B cell (one that has not previously encountered antigen) first encounters the antigen that matches its membranebound antibody, the binding of the antigen to the antibody causes the cell to divide rapidly; its progeny differentiate into memory B cellsand effector B cellscalled plasma cells.
Memory B cells have a longer life span than naive cells, and they express the same membrane-bound antibody as their parent B cell. Plasma cells produce the antibody in a form that can be secreted and have little or no membrane-bound antibody. Although plasma cells live for only a few days, they secrete enormous amounts of antibody during this time. It has been estimated that a single plasma cell can secrete more than 2000 molecules of antibody per second. Secreted antibodies are the major effector molecules of humoral immunity.
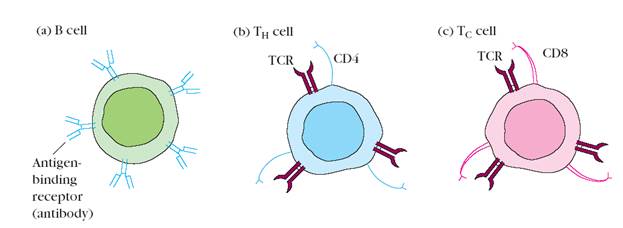
Figure 1. Distinctive membrane molecules on lymphocytes. (a) B cells have about 105 molecules of membrane-bound antibody per cell. All the antibody molecules on a given B cell have the same antigenic specificity and can interact directly with antigen. (b) T cells bearing CD4 (CD4+ cells) recognize only antigen bound to class II MHC molecules. (c) T cells bearing CD8 (CD8+ cells) recognize only antigen associated with class I MHC molecules. In general, CD4+ cells act as helper cells and CD8+ cells act as cytotoxic cells. Both types of T cells express about 105 identical molecules of the antigenbinding T-cell receptor (TCR) per cell, all with the same antigenic specificity.
T LYMPHOCYTES
T lymphocytes also arise in the bone marrow. Unlike B cells, which mature within the bone marrow, T cells migrate to the thymus gland to mature. During its maturation within the thymus, the T cell comes to express a unique antigen-binding molecule, called the T-cell receptor,on its membrane. Unlike membrane-bound antibodies on B cells, which can recognize antigen alone, T-cell receptors can recognize only antigen that is bound to cell-membrane proteins called major histocompatibility complex (MHC) molecules.MHC molecules that function in this recognition event,which is termed “antigen presentation,” are polymorphic (genetically diverse) glycoproteins found on cell membranes. There are two major types of MHC molecules: Class I MHC molecules, which are expressed by nearly all nucleated cells of vertebrate species, consist of a heavy chain linked to a small invariant protein called β2-microglobulin. Class II MHC molecules, which consist of an alpha and a beta glycoprotein chain, are expressed only by antigen-presenting cells. When a naive T cell encounters antigen combined with a MHC molecule on a cell, the T cell proliferates and differentiates into memory T cells and various effector T cells. There are two well-defined subpopulations of T cells: T helper (TH)and T cytotoxic (TC) cells.Although a third type of T cell, called a T suppressor (TS) cell, has been postulated, recent evidence suggests that it may not be distinct from TH and TC subpopulations. T helper and T cytotoxic cells can be distinguished from one another by the presence of either CD4or CD8membrane glycoproteins on their surfaces (Figure 1 b, c). T cells displaying CD4 generally function as TH cells, whereas those displaying CD8 generally function as TC cells.
After a TH cell recognizes and interacts with an antigen–MHC class II molecule complex, the cell is activated – it becomes an effector cell that secretes various growth factors known collectively as cytokines.The secreted cytokines play an important role in activating B cells, TC cells, macrophages, and various other cells that participate in the immune response. Differences in the pattern of cytokines produced by activated TH cells result in different types of immune response.
Under the influence of TH-derived cytokines, a TC cell that recognizes an antigen–MHC class I molecule complex proliferates and differentiates into an effector cell called a cytotoxic T lymphocyte (CTL).In contrast to the TC cell, the CTL generally does not secrete many cytokines and instead exhibits cell-killing or cytotoxic activity. The CTL has a vital function in monitoring the cells of the body and eliminating any that display antigen, such as virus-infected cells, tumor cells, and cells of a foreign tissue graft. Cells that display foreign antigen complexed with a class I MHC molecule are called altered self-cells; these are targets of CTLs.
ANTIGEN-PRESENTING CELLS
Activation of both the humoral and cell-mediated branches of the immune system requires cytokines produced by TH cells. It is essential that activation of TH cells themselves be carefully regulated, because an inappropriate T-cell response to self-components can have fatal autoimmune consequences. To ensure carefully regulated activation of TH cells, they can recognize only antigen that is displayed together with class MHC II molecules on the surface of antigen-presenting cells (APCs). These specialized cells, which include macrophages, B lymphocytes, and dendritic cells, are distinguished by two properties: (1) they express class II MHC molecules on their membranes, and (2) they are able to deliver a co-stimulatory signal that is necessary for TH-cell activation.
Antigen-presenting cells first internalize antigen, either by phagocytosis or by endocytosis, and then display a part of that antigen on their membrane bound to a class II MHC molecule. The TH cell recognizes and interacts with the antigen–class II MHC molecule complex on the membrane of the antigen-presenting cell. An additional costimulatory signal is then produced by the antigen-presenting cell, leading to activation of the TH cell.
Дата добавления: 2016-07-18; просмотров: 2630;
Поиск по сайту
Узнать еще
- Acquired immune deficiency syndrome (AIDS)
- ADAPTIVE RESPONSES IN DISEASE
- Antigen Selection of Lymphocytes Causes Clonal Expansion
- Bacteria Are Simple Cells
- BASIC TUNNELING SYSTEM
- Borderline between compound words and free word-groups. Inseparability of compound words.
- Boundary cases between derivation, inflection and composition.
- CARDIOVASCULAR SYSTEM

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
