Конспект по цитологии
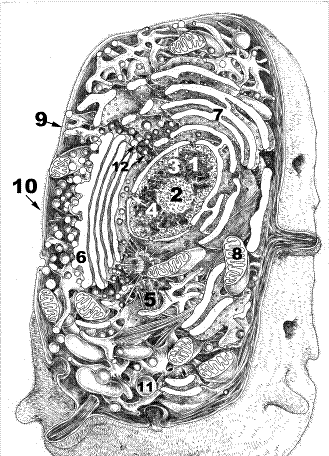 Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами.
Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами.
Рис.1. Объемная реконструкция клетки.
1.Ядро. 2.Ядрышко. 3.Кариолемма. 4.Ядерная пора. 5.Клеточный центр. 6.Комплекс Гольджи. 7.Эндоплазматическая сеть (ЭПС). 8.Митохондрия. 9.Плазмолемма. 10. Гликокаликс. 11.Лизосомы 12.Пероксисомы.
Клетка представляет наименьшую структурно-функциональную единицу живых организмов, способную к самообновлению, саморегуляции и самовоспроизведению. Компоненты клетки распределены в ней упорядоченным образом. Выполнение этого условия обеспечивают мембраны. Мембранные структуры обособляют различные компартменты внутри клетки. Клетка – комплекс компартментов, которые делят её на функциональные зоны. Между структурными элементами клетки происходит взаимодействие благодаря движению гиалоплазмы (циклоз). Составными частями клетки являются её оболочка (плазмолемма), цитоплазма и ядро. Оболочка клетки (9) обеспечивает взаимодействие с другими клетками, с межклеточной жидкостью или другой окружающей средой. Плазмолемма (9) выполняет барьерную, транспортную, рецепторную функции. Толщина плазмолеммы около 10 нм. Она состоит из билипидного слоя в который погружены молекулы белка, которые выполняют транспортную функцию (белки – переносчики), ферментативную, являются циторецепторами. С молекулами плазмолеммы связан гликокаликс (10). Это цепочки полисахаридов, гликолипидов, гликопротеинов. Молекулы гликокаликса участвуют в сцеплении (контактах) клеток и в связывании сигнальных молекул, регулирующих деятельность клетки (гормоны, медиаторы и другие).
Основной объем клетки занят гиалоплазмой (от греч. hyalos-стекло). Это коллоид из воды, ионов и различных молекул органических веществ. В гиалоплазме находятся органеллы и включения. Органеллы – это структурные элементы, выполняющие необходимые для клетки функции. Органеллы, присущие всем клеткам, относят к органеллам общего назначения. Среди них различают органеллы мембранные и немембранные. В приведенном рисунке – схеме отражены мембранные органеллы: гладкая и гранулярная ЭПС (7), комплекс Гольджи (6), лизосомы (11), митохондрии (8), пероксисомы (12). Каждая из указанных органелл имеет на своих мембранах ферменты, обеспечивающие специфику их деятельности.
Немембранные органеллы: клеточный центр, рибосомы, элементы цитоскелета (микротрубочки и промежуточные филаменты), а также актиновые и миозиновые микрофиламенты. Клеточный центр (5) имеет две центриоли (диплосомы), расположенные под углом друг к другу. Каждая центриоль – цилиндр, стенка которого окружена 9 триплетами микротрубочек длиной около 0,5 мкм. Выделяют дочернюю и материнскую центриоль. Вокруг последней формируется центросфера из бесструктурного тонковолокнистого матрикса в который входят концы микротрубочек. Собранные микротрубочки участвуют в структурах цитоскелета. Центриоли в делящихся клетках формируют веретено деления.
Рибосомы обеспечивают синтез белков, состоят из двух субъединиц – большой и малой. Каждая субъединица – комплекс рибосомальной РНК с белками. Большая субъединица содержит 3 молекулы рРНК, малая – одну. Число рибосом в клетке может достигать нескольких миллионов. Те рибосомы, которые не связаны с мембранами ЭПС (свободные рибосомы) синтезируют белки для жизни самой клетки. Рибосомы гранулярной ЭПС (7) синтезируют белки для выведения из клетки. В ядрышке образуются рибосомы, которые через ядерные поры (4) мигрируют в цитоплазму клеток.
Ядро составная часть клетки, содержит кариолемму (3), ядрышко (2), кариоплазму. Размеры ядер могут варьировать. В ядре молекулы ДНК ассоциированы с белками – гистонами. Каждая хромосома образована такими молекулами. Хромосомы – важнейшие структуры, обеспечивающие через синтез ферментов метаболизм клеток.

Назовите структуры, обозначенные цифрами и расскажите об их функциях.
Рис. 2. Схема строения плазмолеммы.
1.Гидрофильные головки фосфолипидов плазмолеммы. 2.Гидрофобные хвосты фосфолипидов. 3.Микротрубочка. 4.Микрофиламент (промежуточный). 5.Интегральные белки. 6.Поверхностный белок. 7.Гликокаликс. 8.Наружный слой липидной мембраны 9.Внутренний слой липидной мембраны. 10.Полуинтегральные белки.
Клеточная мембрана (цитолемма, плазмолемма) отграничивает клетку от внешней среды, осуществляет ее взаимодействие с внешней средой, в том числе и с другими клетками, обеспечивает избирательную проницаемость веществ, участвует в поддержании гомеостаза. Согласно современным представлениям, клеточная мембрана, как и остальные мембраны клетки, состоит из двух слоев липидов со встроенными в них белками.
Липиды составляют от 25 до 40% массы. Это, в основном, фосфолипиды, сфинголипиды и холестерин. Их полярная часть (головка) является гидрофильной (1) и располагается поверхностно. Гидрофобная же часть (2) погружена внутрь мембраны. Белки составляют около 60% массы и обеспечивают специализированные функции мембраны. Это глобулярные белки. Они формируют скопления в плазмолемме. По степени погружения в мембрану белки могут располагаться поверхностно (с наружной или внутренней поверхности) (6), пронизывать всю толщу мембраны – интегральные белки (5); или глубоко погружаться, но не достигать одной из сторон – полуинтегральные белки (10). Интегральные белки играют роль в осуществлении трансмембранных процессов.
С учетом разнообразия функций белки можно весьма условно разделить на группы: рецепторные белки - лежат поверхностно, взаимодействуют с внешними сигнальными молекулами; белки-переносчики - осуществляют трансмембранный перенос; адгезивные белки - обеспечивают прикрепление клетки к другим клеткам или к элементам межклеточного вещества; ферменты; структурные белки, в том числе и осуществляющие опорно-каркасные функции (они чаще всего связаны с микротрубочками, микрофиламентами и филаментами с внутренней стороны мембраны и с адгезивными белками с внешней стороны).
Углеводы содержатся на внешней поверхности клеточной стенки и участвуют в формировании гликокаликса (7). Это, в основном, олигосахариды, связанные с пептидами (гликопротеины) или липидами (гликолипиды). Гликопротеины имеют высокую видовую, индивидуальную и тканевую специфичность.
В подмембранные комплексы входят элементы цитоскелета (3,4). Эти элементы могут участвовать в формировании плотных и десмосомальных межклеточных контактов.
Транспортные процессы через мембрану (трансмембранный перенос):
1.Пассивная диффузия по градиенту концентрации. Осуществляется в обе стороны в зависимости от парциального давления (концентрации) того или иного вещества. Не требует затрат энергии.
2. Облегченная диффузия. Отличается от пассивной тем, что возможность пассивной диффузии по градиенту концентрации может осуществляться только в одном направлении (либо в клетку, либо из нее). Облегченная диффузия осуществляется через специальные белки-переносчики, являющиеся интегральными, высокоспециализированными белками, осуществляющими перенос строго определенных веществ, зависящий от функциональной активности клетки. Они могут быть «открытыми», формируя «каналы», через которые проникает вещество, или «закрытыми».
3. Активный транспорт через клетку осуществляется с затратами энергии, против градиента концентрации, высокоспецифичен и нередко сопряжен с транспортом двух веществ в противоположном направлении. Так в клетку попадают ионы калия и одновременно выводятся ионы натрия. Высокомолекулярные вещества, макромолекулярные комплексы могут транспортироваться в клетку или выводиться из нее путем эндоцитоза или экзоцитоза.
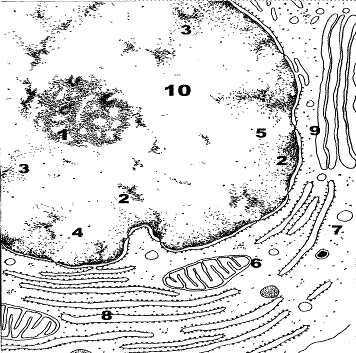
Назовите структуры, обозначенные цифрами. Укажите функции этих структур.
Рис. 3.Ядро и цитоплазма клетки.
1.Ядрышко. 2.Гетерохроматин. 3.Эухроматин. 4.Ядерная оболочка. 5.Ядерная пора. 6.Митохондрия. 7.Лизосома. 8.Гранулярная ЭПС. 9.Комплекс Гольджи. 10.Кариоплазма.
Ядро состоит из кариоплазмы (10) и кариолеммы (4). В кариоплазме, представляющей коллоид в виде геля, можно выделить ядрышко (1), матрикс, хроматин (2,3). В ядре имеется от одного до нескольких ядрышек. Они могут иметь различные размеры, форму, плотность и область распределения в зависимости от функциональной активности клетки. Основная функция ядрышка – синтез рибосомальной РНК (рРНК) и субъединиц рибосом.
Хроматин ядра – комплекс дезоксирибонуклеиновой кислоты с белками, где ДНК находится в различной степени спирализации. По электронной и светооптической плотности выделяют электронно-плотный, грубо окрашенный гетерохроматин (2) и более нежно окрашенный, менее электронно-плотный эухроматин (3). Гетерохроматин- зона сильно конденсированной ДНК. При электронной микроскопии формирует темные глыбки неправильной формы. В гетерохроматине ДНК, в основном, связана с гистоновыми белками. Гетерохроматин представляет собой плотно упакованные скопления нуклеосом. В зависимости от локализации подразделяется на пристеночный, матричный и перинуклеарный. Пристеночный гетерохроматин прилежит к внутренней поверхности ядерной оболочки, матричный – распределен в матриксе кариоплазмы. Как вариант матричного – перинуклеарный гетерохроматин, примыкает к ядрышку. Эухроматин – это область слабо конденсированной ДНК. С нуклеиновыми кислотами в эухроматине связаны, в основном, негистоновые белки (проявляют менее выраженные основные свойства, более разнообразны по химическому составу).
Ядерная оболочка (кариолемма) (4) отграничивает содержимое ядра от цитоплазмы. Представлена двумя мембранами и перинуклеарным пространством между ними (перинуклеарная цистерна). В ней имеются ядерные поры – области слияния наружной и внутренней мембран (5).
Ядерная пора (5) обеспечивает избирательный транспорт макромолекул. Через нее свободно диффундируют по градиенту концентрации молекулы воды, растворенные в ней газы, неорганические ионы, низкомолекулярные органические вещества. Содержимое ядра отличается от цитоплазмы по составу органических веществ высокого молекулярного веса (ферменты, макромолекулярные соединения); по составу низкомолекулярных веществ, приближено к матриксу цитоплазмы. На внутренней и наружной поверхности комплекса ядерной поры имеются высокоспецифичные рецепторы, обеспечивающие транспорт из ядра субъединиц рибосом, информационной РНК (иРНК), транспортной РНК (тРНК) и некоторых других веществ. В ядро же избирательно транспортируются ферменты, белки ламин, гистоны. Транспорт макромолекул происходит активно, а белки комплекса ядерной поры обладают АТФ-азной активностью.
Наружная ядерная мембрана по набору рецепторов и составу аналогична гранулярной или агранулярной ЭПС. Внутренняя ядерная мембрана участвует в формировании ядерной пластинки. Ядерная пластинка структурирует ядерную оболочку и перинуклеарный хроматин. Содержит белки – ламины, которые прикрепляются к белкам внутренней мембраны, выполняющим опорно-каркасные функции. Ламины образуют фибриллярные структуры, аналогичные промежуточным филаментам цитоплазмы. К внутренней поверхности ядерной оболочки нередко прикрепляется гетерохроматин (пристеночный). Перинуклеарная цистерна (пространство) характеризуется низкой электронной плотностью. Состав ферментов аналогичен содержимому гранулярной ЭПС.
Цитоплазма состоит из матрикса (гиалоплазма), органелл и включений. Гиалоплазма может иметь свойства золя или геля. Органеллы – постоянно присутствующие структурные элементы цитоплазмы. Из органелл представленных на рисунке: митохондрии (6), лизосомы (7), гр. ЭПС (8), комплекс Гольджи (9).
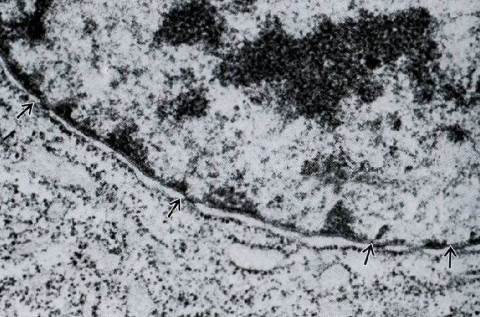
Назовите структуры клетки, изображенные на электронной микрофотографии. Укажите тип электронной микроскопии.
Рис. 4. Фрагмент клетки с участком ядра. Видна ядерная оболочка (кариолемма), состоящая из двух мембран – внутренней и наружной. Стрелками показаны ядерные поры. Внутри ядра виден электронно-плотный гетерохроматин и мелкозернистый эухроматин. В цитоплазме вокруг ядра находится гранулярная ЭПС. Трансмиссионная электронная микроскопия. Ув. 50000.

Назовите структуры клетки, изображенные на электронной микрофотографии. Укажите тип электронной микроскопии.
Рис. 5. Фрагмент клетки с участком ядра. Видна кариолемма с ядерными порами (стрелка) и диктиосомы комплекса Гольджи. Сканирующая электронная микроскопия, метод замораживания-скалывания-травления, позволяющий изучить строение клеточных мембран. Ув. 22000.

Назовите структуры, обозначенные цифрами. Расскажите о последовательности процессов в ходе взаимодействия гранулярной ЭПС и комплекса Гольджи.
Рис.6.Схема взаимодействия гранулярной эндоплазматической сети и комплекса Гольджи.
1.Гранулярная эндоплазматическая сеть (ЭПС). 2.Гладкая эндоплазматическая сеть (ЭПС). 3.Комплекс Гольджи. 4.Диктиосомы. 5.Транспортные пузырьки (от ЭПС). 6.Лизосома.
До недавнего времени не были раскрыты механизмы взаимодействия эндоплазматической сети и комплекса Гольджи. В настоящее время доказано наличие конвейера в воспроизводстве веществ в клетке. Первым блоком этого конвейера является эндоплазматическая сеть, а вторым комплекс Гольджи. Эти блоки взаимосвязаны.
Эндоплазматическая сеть представлена трубочками, цистернами, мембранами, пузырьками. Большинство веществ синтезируется на наружной поверхности мембран. Затем вещества переносятся внутрь каналов эндоплазматической сети. На конце каналов эндоплазматической сети вещества накапливаются, а затем отделяются в виде транспортных пузырьков (5). Если образуются вещества для обновления мембран, то сборка мембран происходит в комплексе Гольджи, там же образуются лизосомы (6). Комплекс Гольджи является местом конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном, в эндоплазматической сети. При электронной микроскопии комплекс Гольджи состоит из скоплений плоских цистерн (диктиосом)(4) и пузырьков. Наиболее широкие уплощенные цистерны обращены в сторону эндоплазматической сети. Транспортные пузырьки от эндоплазматической сети переходят к цистернам (цис-полюс) комплекса Гольджи и там происходит их дальнейшая трансформация: образуются комплексы белков с углеводами и липидами. Образованные вещества переходят из одних цистерн в другие. На белковых концах цистерн (транс-полюс) образуются выпячивания, которые отщепляются в виде пузырьков и направляются в гиалоплазму или к плазмолемме для выведения, а в комплекс Гольджи поступают новые порции продуктов, синтезированных в гранулярной эндоплазматической сети.
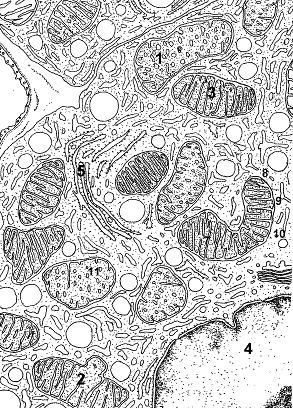
Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами.
Рис. 7. Фрагмент клетки. Митохондрии.
1.Крупная митохондрия с трубчатыми кристами. 2.Деление митохондрии путем почкования. 3.Митохондрии овальной формы с трабекулярными кристами. 4.Ядро. 5.Гранулярная эндоплазматическая сеть. 6.Трубчатая криста. 7.Трабекулярная криста. 8.Наружная мембрана. 9.Внутренняя мембрана. 10.Перимитохондриальное пространство. 11.Матрикс митохондрии.
Митохондрии имеют средние поперечные размеры 0,5 – 3 мкм. В зависимости от размеров можно выделить мелкие, средние, крупные, гигантские органеллы. По форме они могут быть округлые, овальные (3) и нитевидные. Митохондрии состоят из матрикса (11); внутренней мембраны (9), образующей кристы (6,7) (впячивания в матрикс); перимитохондриального пространства (10) и наружной мембраны (8). Форма крист может быть в виде пластинок (7)(трабекулярные) и трубочек (6) (мультивезикулярные).
Матрикс (11) митохондрии гомогенный, иногда мелкозернистый, различной электронной плотности. В матриксе содержатся ферменты, одноцепочечная циклическая ДНК, митохондриальная РНК, митохондриальные рибосомы, высокая концентрация ионов Са2+. ДНК митохондрий близка к ДНК прокариот. По строению митохондриальные рибосомы близки к рибосомам прокариотических организмов, но меньшего размера.
Внутренняя мембрана митохондрии (9) обладает избирательной проницаемостью для электронов, образующихся при аэробном фосфорилировании и слабопроницаема для других веществ. К ее внутренней поверхности прикрепляются тесно прилежащие друг к другу ферменты цепи окислительного фосфорилирования.
Наружная мембрана митохондрии (8) более проницаема для низкомолекулярных веществ. На поверхности наружной мембраны имеются специфические рецепторные комплексы, через которые белки из матрикса могут транспортироваться в перимитохондриальное пространство, или, за счет временного слияния с внутренней мембраной, попасть в матрикс митохондрии.
В митохондриях имеются два автономных компартмента. Перимитохондриальное пространство (внешний компартмент) формируется за счет проникновения белковых комплексов гиалоплазмы, и внутренний компартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК.
Во внутреннем компартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са2+ в сравнении с гиалоплазмой. Ферментная активность внешнего и внутреннего компартментов, состав белков также сильно различаются. Внутренний компартмент нередко имеет более высокую электронную плотность, чем внешний.
Специфическими маркерами митохондрии являются ферменты цитохромоксидаза и сукцинатдегидрогеназа.
Некоторые авторы считают, что в клетках не множество, а одна гигантская митохондрия, которая многократно попадает в срез препарата, однако большинство исследователей указывает на ошибочность данной концепции, во всяком случае, для большинства клеток организма человека. Новые митохондрии образуются путем деления, способом перетяжки или почкования (2). В последнем случае образуется протомитохондрия, постепенно увеличивающаяся в размерах. При размножении в каждой из вновь образующихся митохондрий имеется свой собственный геном.
Старые митохондрии разрушаются путем аутолиза (самопереваривания клеткой с помощью лизосом) с образованием аутолизосом. Из аутолизосомы образуется остаточное тельце. При полном переваривании содержимое остаточного тельца выводится путем экзоцитоза. При неполном переваривании остатки митохондрий могут накапливаться в клетке в виде слоистых телец или гранул с липофусцином. Часть митохондрий может обызвествляться с образованием нерастворимых солей кальция (кальцинатов).
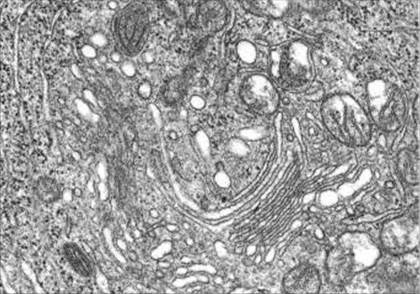
Какие органоиды показаны на электронной микрофотографии? Укажите тип электронной микроскопии.
Рис. 8. Фрагмент клетки.Вцентре расположен комплекс Гольджи, состоящий из диктиосом, также в цитоплазме видны митохондрии и гранулярная ЭПС. Трансмиссионная электронная микроскопия. Ув. 30000.
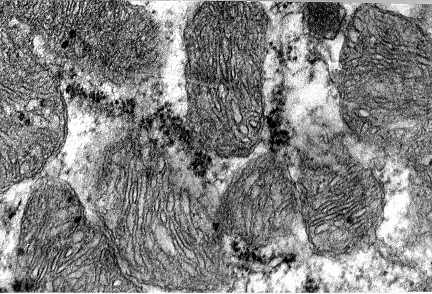
Какие органоиды показаны на электронной микрофотографии? Укажите тип электронной микроскопии.
Рис. 9. Митохондрии в эпителиоцитах тонкой кишки. Видны наружная и внутренняя мембраны митохондрий, пластинчатые (ламеллярные) кристы. Трансмиссионная электронная микроскопия. Ув. 20000.
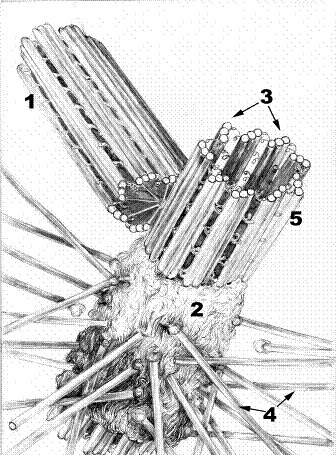
Назовите структуры, обозначенные цифрами, укажите их функции.
Рис.Клеточный центр.
1.Дочерняя центриоль. 2. Тонковолокнистый матрикс. 3. Триплет из микротрубочек 4. Микротрубочки центросферы. 5. Материнская центриоль. 6.Ручки с динеинами.
Клеточный центр образован двумя перпендикулярно расположенными центриолями (1) и центросферой. Центриоли имеют цилиндрическую форму. Их поперечный диаметр составляет около 150 нм, а длина – 500 нм. Выделяют дочернюю и материнскую центриоли (5). Вокруг материнской центриоли формируется центросфера (4) из бесструктурного тонковолокнистого матрикса (2), в который входят концы микротрубочек.
Центриоли составлены триплетами микротрубочек (3). Соседние триплеты взаимодействуют между собой с помощью ручек, образованных молекулами белка динеина (6). Динеин обладает АТФ-азной активностью и способен вызвать смещение (скольжение) одного триплета микротрубочек по отношению к другому, обеспечивая движение.
Клеточный центр в интерфазной клетке организует ее компартменты, участвует в циклозе. При митотическом и мейотическом делении в животной клетке центриоли обеспечивают равномерное расхождение генетического и цитоплазматического материала делящейся клетки, участвуют в формировании веретена деления.
Во время деления центриоли расходятся к полюсам, формируя новые клеточные центры клеток. Центриоли способны индуцировать синтез новых центриолей, до нескольких десятков или даже сотен на одну клетку. Вновь образуемые центриоли перемещаются на периферию и служат основой для формирования ресничек, формируя в них базальное тельце.
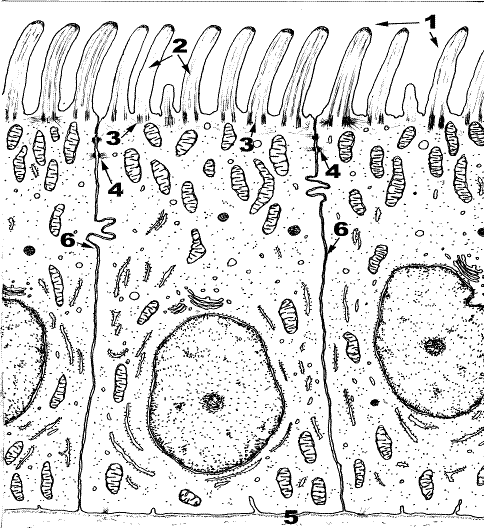
Назовите структуры, обозначенные цифрами, объясните функции этих структур.
Рис. 11. Специальные структуры поверхности клетки. Реснички в эпителиоцитах.
1. Ресничка. 2.Аксонема. 3.Базальное тельце. 4.Десмосома. 5.Базальная мембрана. 6. Плазмолемма.
Реснички относят к специализированным органоидам клетки. Они присутствуют в эпителии воздухоносных путей, маточных труб и обладают подвижностью.
Ресничка (1) представляет собой выпячивание цитоплазмы, окруженное клеточной мембраной (6) диаметром 300 нм. В основании ресничек мелкие гранулы – базальные тельца (3), которые погружены в цитоплазму. К базальному тельцу могут прикрепляться спутники от которых отходят микротрубочки. Внутри реснички находится аксонема («осевая нить») (2), выступающая в просвет или полость органа, выстланного реснитчатым эпителием. Аксонема состоит из микротрубочек. Микротрубочки собраны в комплексы в виде дуплетов: по 9 пар на периферии и одна пара в центре. Каждая из двух первых микротрубочек девяти триплетов базального тельца является основой для дуплетов микротрубочек цитоплазматического выроста. Таким образом, две микротрубочки триплетов базального тельца являются микротрубочками дуплетов аксонемы. Соседние дуплеты соединяются друг с другом с помощью «ручек» из белка динеина, который обладает АТФ-азной активностью. В присутствии ионов кальция динеины сокращаются, потребляя при этом энергию, и микротрубочки скользят относительно друг друга. Все микротрубочки достигают своим дистальным отрицательным полюсом гомогенный матрикс аксонемы. Таким образом, базальное тельце и аксонема структурно связаны и составляют единое целое. Основной белок ресничек – тубулин к сокращению не способен. Смещения дуплетов микротрубочек относительно друг друга вызывают изгиб всей реснички. Движение микроресничек обеспечивает белок динеин. Реснички образуются за счет центриолей: от материнской синтезируется дочерняя. Вновь синтезированная органелла смещается к периферии. Процесс многократно повторяется и под мембраной образуется много базальных телец, которые являются основой для образования ресничек. Движения ресничек зависят от содержания внутриклеточного кальция, АТФ, ионного состава межклеточного вещества и т.д.
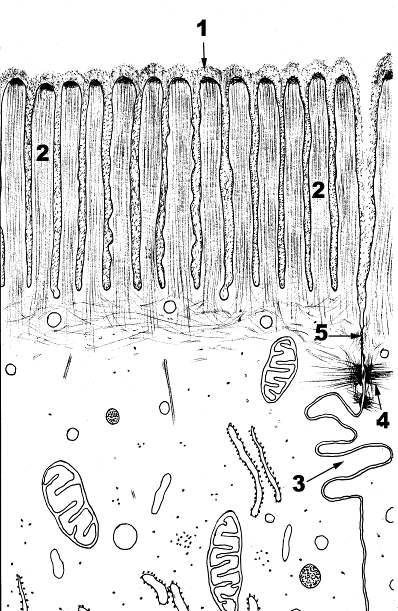
Назовите структуры, обозначенные цифрами, укажите функции этих структур.
Рис. 12. Специальные структуры поверхности клетки: микроворсинки в эпителиоцитах
1.Гликокаликс. 2.Микроворсинки. 3.Межклеточный контакт в виде «замка». 4.Десмосома. 5. Плотный контакт
Микроворсинки (2) видны под световым микроскопом как тонкие выросты клетки. За счет микроворсинок увеличивается площадь взаимодействия клетки с внешней средой. Микроворсинки состоят из клеточной мембраны, гиалоплазмы и тонких микрофиламентов (актиновых). Последние располагаются параллельно поверхности мембраны в виде упорядоченных пучков. Положительный полюс направлен к периферии и связывается с электронноплотным веществом «верхушки» микроворсинки. В основании микроворсинки актиновые филаменты вплетаются в сеть подобных структур, которые образуют кутикулу. С мембранами тонкие (актиновые) филаменты взаимодействуют с помощью минимиозина и виллина. Взаимодействие с минимиозином способствует сокращению микроворсинки. Микроворсинки являются неотьемлемой частью эпителиоцитов тонкой и толстой кишки, канальцев почки. В тонкой кишке ферменты, прикрепленные к гликокаликсу (1) и находящиеся в толще билипидного слоя мембраны микроворсинок, обеспечивают всасывание, переваривание веществ на поверхности клеток. Большое количество микроворсинок в клетке создает между ними узкие щели. В этих пространствах действуют капиллярные силы, способствующие присасыванию жидкости. Величина и размеры микроворсинок непостоянны. В почках микроворсинки клеток канальцев нефрона всасывают воду и электролиты, последние переходят в кровь.
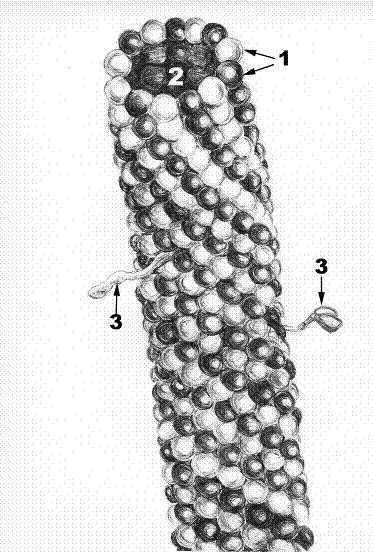
Назовите элементы микротрубочки. Назовите структуры, в которых есть микротрубочки. Опишите варианты функций микротрубочек.
Рис. 13.Схема строения микротрубочки.
1. Димер микротрубочки. 2. Просвет микротрубочки. 3. Ассоциированные белки.
Микротрубочки представляют собой полые цилиндры диаметром 20 – 30 нм, внутренний просвет имеет ширину 15 нм (2). Являются элементами цитоскелета. Микротрубочки эволюционно древние образования клеток. Они участвуют в делении клеток, входят в состав центриолей, микроресничек, цитоскелета. В клетках микротрубочки обеспечивают внутриклеточный транспорт, в нейронах участвуют в аксотоке. Микротрубочки могут быть разбросаны в цитоплазме, либо собраны в виде пучков, например, в составе митотического веретена, а могут сливаясь друг с другом формировать пары ( в жгутиках), триплеты (в центриолях).
Микротрубочки состоят из глобулярных белков – тубулинов, на поперечном срезе их 13. Тубулины подразделяются на альфа и бета тубулины. Альфа тубулины соединяются в виде цепочки, которая формирует спираль. Одновременно, в виде параллельной спирали полимеризуются бета-тубулины, т.е. образуются димеры тубулина (1). Таким образом, тубулины могут быть в двух формах – глобулярной (диспергированной в матриксе) и фибриллярной – в виде микротрубочек. С поверхностью микротрубочки могут быть связаны ассоциированные белки (3). Если эти белки сократительные (динеин), то их сокращение приводит к изменению положения и формы микротрубочки. Микротрубочки имеют полярность. У них есть положительный (+) и отрицательный (-) концы. Со стороны (-) полюса, лежащего ближе к клеточному центру, тубулины полимеризуются медленнее и легко распадаются до глобулярных частиц. Со стороны (+) конца легче осуществляется полимеризация и он направлен на периферию клетки.
Воздействие на микротрубочки может привести к нарушению их функций и реконструкции, к их разрушению. Это используют в лечении больных со злокачественными опухолями, при этом нарушается деление злокачественных клеток.
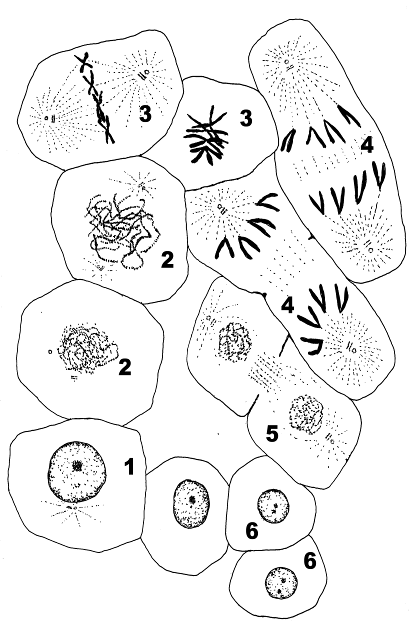
Дайте название процессу и его фазам. Опишите изменения, происходящие на каждой из приведенных схем.
Рис.14.Митоз.
1. Клетка в интерфазе. 2, 3 Профаза. 4, 5. Метафаза. 6, 7.Анафаза. 8.Телофаза. 9.Дочерние клетки.
В предмитотической фазе (1) в клетке удваивается число хромосом. В профазе (2,3) прекращается транскрипция, начинается спирализация хромосом (стадия рыхлого клубка), исчезает ядрышко. Распадается оболочка ядра. К концу профазы видны хромосомы, каждая из которых состоит из двух хроматид. Хроматиды сплетены и не видны отдельно. Характерной чертой профазы является формирование веретена деления. К каждому полюсу отходят по две центриоли и от них образуются микротрубочки. Формирование микротрубочек обеспечивает полимеризация белков тубулинов. С микротрубочками связываются хромосомы.
Метафаза (4, 5) занимает 20-30 минут. В этот период завершено образование веретена деления и хромосомы распределяются в области экватора. К концу метафазы сестринские хроматиды разделяются.
В анафазе (6,7) сестринские хроматиды становятся самостоятельными хромосомами и расходятся к полюсам формирующихся клеток. Это происходит при участии микротрубочек. Телофаза (8) является окончанием митоза. Ранняя телофаза – это завершение расхождения хромосом. В поздней телофазе начинается формирование новых ядер, обособление генетического материала. Завершается поздняя телефаза разделением исходной клетки на две дочерние (9) (цитокинез или цитотомия). Хромосомы начинают транскрибировать РНК. К концу телофазы ядрышко полностью сформировано.
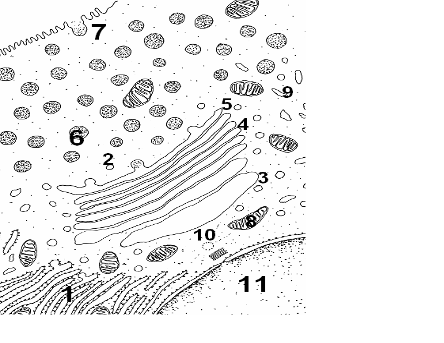 Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами.
Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами.
Рис. 15. Фрагмент клетки. Процессы синтеза, накопления и выделения секрета.
1.Гранулярная ЭПС. 2.Транспортные пузырьки. 3.Цис-компартмент комплекса Гольджи. 4.Промежуточный компартмент комплекса Гольджи. 5.Транс-компартмент комплекса Гольджи 6.Секреторные включения. 7.Секреция (экзоцитоз). 8.Митохондрия. 9.Агранулярная ЭПС. 10.Клеточный центр. 11.Ядро.
Местом конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки (в основном, в эндоплазматической сети) является комплекс Гольджи. Под электронным микросокпом органелла представляет собой скопления плоских цистерн, которые называются диктиосомы. Таких скоплений в клетке может быть несколько. Цистерны плотно прилежат друг к другу. В центральной части просвет между ними невелик, а на периферии цистерны образуют расширения – ампулы, ширина которых непостоянна. В каждой стопке около десяти цистерн. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул). Их больше по краям комплекса Гольджи. Одни авторы полагают, что цистерны взаимодействуют между собой за счет транспортных пузырьков (2), другие – что цистерны имеют участки слияния. Комплекс Гольджи поляризован, т.е. качественно не однороден с разных сторон и состоит в свою очередь из нескольких компартментов. Он имеет незрелую цис-поверхность, лежащую ближе к ядру (11), и зрелую транс-поверхность, обращенную к наружной мембране клетки. Таким образом, органелла состоит из нескольких компартментов, которые, выполняя специфические функции, функционально взаимосвязаны между собой. Цис-компартмент (3) обычно обращен к клеточному центру (10). Его внешняя поверхность обычно имеет выпуклую форму. С его цистернами сливаются микровезикулы (транспортные пиноцитозные пузырьки), направляющиеся из эндоплазматической сети. Промежуточный компартмент (4) составлен несколькими цистернами в центре органеллы. Осуществляет гликозилирование (присоединяет углеводные цепочки), фосфорилирование (присоединение фосфорных остатков), карбоксилирование (присоединение карбоксильных групп), синтез гликолипидов и липопротеинов. В нем, как и в цис-компартменте, происходит формирование третичных и четвертичных белковых комплексов. Транс-компартмент (5) располагается ближе к периферии клетки. Внешняя поверхность обычно вогнутая. Частично он переходит в транс-сеть – систему везикул, вакуолей и канальцев. Отдельные диктиосомы могут быть связаны друг с другом системой пузырьков и цистерн, примыкающих к дистальному концу скопления плоских мешков, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах (транс-сеть комплекса Гольджи). В структурах транс-компартмента происходит сортировка (сегрегация) белков и других веществ, образование секреторных гранул, предшественников первичных лизосом и пузырьков спонтанной секреции. Секреторные пузырьки и прелизосомы являются окаймленными и окружены белком – клатрином. Окаймленные пузырьки отщепляются от транс-сети и их перемещение является гормонозависимым. Процесс транспортировки окаймленных пузырьков находится под влиянием микротрубочек. Белковые комплексы вокруг пузырьков распадаются после отщепления пузырька от транс-сети и вновь формируются в момент секреции. Пузырьки спонтанной секреции не окружены клатрином, их формирование происходит непрерывно и они, направляясь к клеточной мембране, сливаются с ней, обеспечивая ее восстановление.
Таким образом, в комплексе Гольджи происходит формирование сложных биополимерных комплексов и, в частности, гликозилирование белков и липидов, в результате чего образуются гликопротеины, гликолипиды. В органелле формируются третичные и четвертичные структуры белков, частично происходит созревание секрета, его конденсация и упаковка.
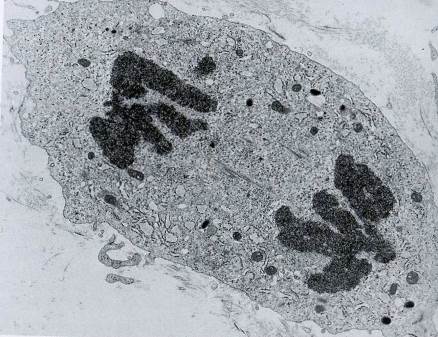
Какой процесс показан на электронной микрофотографии? Назовите стадию процесса, аргументируя вывод. Укажите тип электронной микроскопии.
Рис. 16. Митотическое деление клетки, анафаза. Видны дочерние наборы хромосом, расходящиеся к полюсам клетки. Трансмиссионная электронная микроскопия. Ув. 1400.
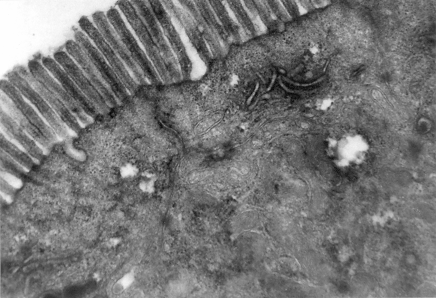
Какие структуры изображены на электронной микрофотографии, каково их значение для клетки? Укажите тип электронной микроскопии.
Рис. 17. Микроворсинки эпителиальных клеток тонкой кишки.Ониувеличивают площадь всасывающей поверхности клетки, в центре микроворсинок видны пучки микрофиламентов. Трансмиссионная электронная микроскопия. Ув. 5000.
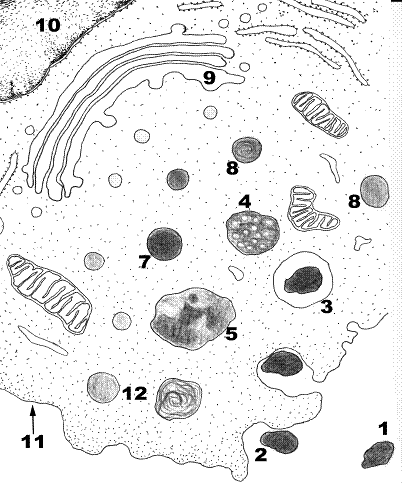
Назовите структуры, обозначенные цифрами. Опишите сущность процессов фагоцитоза и пиноцитоза.
Рис. 18. Фагоцитоз и пиноцитоз.
1.Макромолекулярная структура. 2.Начало фагоцитоза. 3.Фагосома. 4.Фаголизосома (вторичная лизосома). 5.Гранулярная ЭПС. 6.Митохондрия. 7.Первичная лизосома. 8.Эндосома. 9. Аппарат Гольджи. 10.Фрагмент ядра. 11.Плазмолемма. 12.Остаточное тельце.
Существует несколько способов эндоцитоза (от греч. endon – внутри, kytos – клетка: пиноцитоз (от греч. pino – пью) и фагоцитоз (от греч. phagos – пожирающий). При пиноцитозе клетка захватывает жидкие коллоидные частицы, а при фагоцитозе – плотные частицы (макромолекулярные комплексы, части клеток, бактерии и др.). В этих процессах активную роль играют плазмолемма (11) и гликокаликс.
При пиноцитозе и фагоцитозе вбираемые клеткой частицы взаимодействуют с плазмолеммой и окружаются ею (2). Жидкие частицы окаймляются дополнительно белком клатрином (окаймленные пузырьки) (12). В последующем и при пиноцитозе и при фагоцитозе происходит взаимодействие захваченных клеткой веществ с лизосомами.
Фагоцитоз характерен для клеток макрофагов, которые находятся в рыхлой соединительной ткани в каждом органе, нейтрофилов и др. При фагоцитозе бактерий, частей клеток специализированными клетками идет взаимодействие фагоцитируемой частицы с рецепторами на поверхности клетки и активация фагоцитоза с изменением внутриклеточного содержания кальция. Это ведет к изменению полимеризации тонких микрофиламентов и микротрубочек, вызывает формирование выпячивания цитолеммы (псевдоподии) с погружением крупной частицы внутрь клетки (образование фагосомы). В дальнейшем эндоцитозные пузырьки (эндосомы) (8) могут сливаться друг с другом и внутри пузырьков, кроме поглощенных веществ обнаруживаются гидролитические ферменты, которые поступают из лизосом. Ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиалоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению.
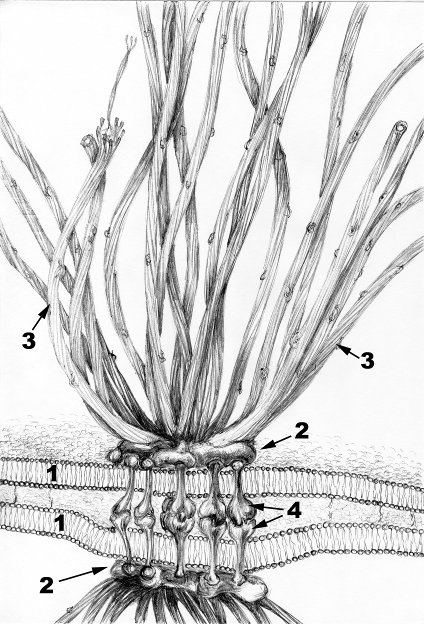
Назовите структуру, приведенную на фотограмме. Укажите детали, обозначенные цифрами и их значение.
Рис.19.Структура десмосомы.
1. Плазмолеммы контактирующих клеток 2. Пятно десмосомы 3. Промежуточные филамен
| <== предыдущая лекция | | | следующая лекция ==> |
| ОРГАНЫ ДЫХАТЕЛЬНОЙ СИСТЕМЫ | | | Механическое движение. Относительность движения. Равномерное и равноускоренное движение. |
Дата добавления: 2016-07-18; просмотров: 4599;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
