Защитная (амортизационная)
Толстый слой жира защищает внутренние органы многих животных от повреждений при ударах (например, сивучи при массе до тонны, могут прыгать в воду со скал высотой 20-25 м).
Увеличения плавучести
Самые разные организмы — от диатомовых водорослей до акул — используют резервные запасы жира как средство снижения среднего удельного веса тела и, таким образом, увеличения плавучести. Это позволяет снизить расходы энергии на удержание в толще воды.
Классификация липидов
Классификация липидов, как и других соединений биологической природы, — весьма спорный и проблематичный процесс. Предлагаемая ниже классификация хоть и широко распространена в липидологии, но является далеко не единственной. Она основывается, прежде всего, на структурных и биосинтетических особенностях разных групп липидов.
Простые липиды — липиды, включающие в свою структуру углерод (С), водород (H) и кислород (O).
- Жирные кислоты — алифатические одноосновные карбоновые кислоты с открытой цепью, содержащиеся в этерифицированной форме в жирах, маслах и восках растительного и животного происхождения.
- Жирные альдегиды — высокомолекулярные альдегиды, с числом атомов углерода в молекуле выше 12.
- Жирные спирты — высокомолекулярные спирты, содержащие 1-3 гидроксильные группы
- Предельные углеводороды с длинной алифатической цепочкой
- Сфингозиновые основания
- Воски — сложные эфиры высших жирных кислот и высших высокомолекулярных спиртов.
- Триглицериды (Жиры)
Сложные липиды — липиды, включающие в свою структуру помимо углерода(С), водорода(H) и кислорода(О) другие химические элементы. Чаще всего: фосфор(Р), серу(S), азот(N).
- Полярные
- Фосфолипиды — сложные эфиры многоатомных спиртов и высших жирных кислот, содержащие остаток фосфорной кислоты и соединённую с ней добавочную группу атомов различной химической природы.
- Гликолипиды — сложные липиды, образующиеся в результате соединения липидов с углеводами.
- Фосфогликолипиды
- Сфинголипиды — класс липидов, относящихся к производным алифатических аминоспиртов.
- Мышьяколипиды
- Нейтральные
- Ацилглицериды
- Диглицериды
- Моноглицериды
- Триглицериды
- Церамиды
- Эфиры стеринов
- N-ацетилэтаноламиды
Оксилипиды
- Оксилипиды липоксигеназного пути
- Оксилипиды циклооксигеназного пути
Молекулы простых липидов состоят из спирта, жирных кислот, сложных — из спирта, высокомолекулярных жирных кислот, возможны остатки фосфорной кислоты, углеводов, азотистых оснований и др. Строение липидов зависит в первую очередь от пути их биосинтеза.
Аминокислоты
Аминокисло́ты (аминокарбо́новые кисло́ты) — органические соединения, в молекуле которых одновременно содержатся карбоксильные и аминные группы.
Аминокислоты могут рассматриваться как производные карбоновых кислот, в которых один или несколько атомов водорода заменены на аминные группы.
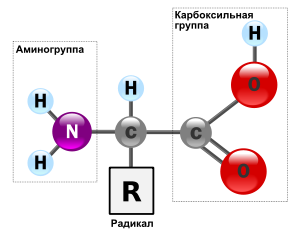
Все аминокислоты — амфотерные соединения, они могут проявлять как кислотные свойства, обусловленные наличием в их молекулах карбоксильной группы —СООН, так и основные свойства, обусловленные аминогруппой —NH2.
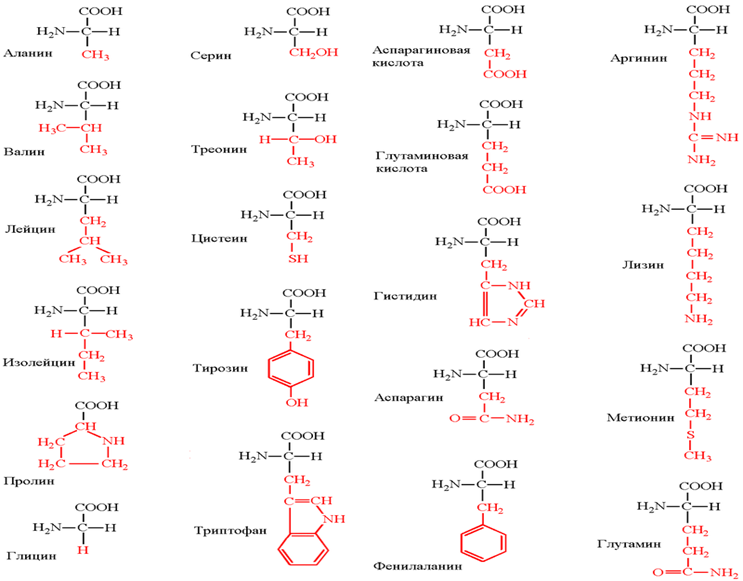
В процессе биосинтеза белка в полипептидную цепь включаются 20 α-аминокислот, кодируемых генетическим кодом. Помимо этих аминокислот, называемых протеиногенными, или стандартными, в некоторых белках присутствуют специфические нестандартные аминокислоты, возникающие из стандартных в процессе посттрансляционных модификаций. В последнее время к протеиногенным аминокислотам иногда причисляют трансляционно включаемые селеноцистеин (Sec, U) и пирролизин (Pyl, O). Это так называемые 21-я и 22-я аминокислоты.
| Глицин | Gly | G | Glycine |
| Аланин | Ala | A | Alanine |
| Валин | Val | V | Valine |
| Изолейцин | Ile | I | Isoleucine |
| Лейцин | Leu | L | Leucine |
| Пролин | Pro | P | Proline |
| Серин | Ser | S | Serine |
| Треонин | Thr | T | Threonine |
| Цистеин | Cys | C | Cysteine |
| Метионин | Met | M | Methionine |
| Аспарагиновая кислота | Asp | D | asparDic acid |
| Аспарагин | Asn | N | asparagiNe |
| Глутаминовая кислота | Glu | E | gluEtamic acid |
| Глутамин | Gln | Q | Q-tamine |
| Лизин | Lys | K | before L |
| Аргинин | Arg | R | aRginine |
| Гистидин | His | H | Histidine |
| Фенилаланин | Phe | F | Fenylalanine |
| Тирозин | Tyr | Y | tYrosine |
| Триптофан | Trp | W | tWo rings |
Незаменимые аминокислоты — необходимые аминокислоты, которые не могут быть синтезированы в том или ином организме, в частности, в организме человека. Поэтому их поступление в организм с пищей необходимо.
Незаменимыми для взрослого здорового человека являются 8 аминокислот: валин, изолейцин, лейцин, лизин, метионин, треони́н, триптофан и фенилалани́н;
Для детей незаменимыми также являются аргинин и гистидин.
Белки
Белки́ (протеи́ны, полипепти́ды) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью. В живых организмах аминокислотный состав белков определяется генетическим кодом, при синтезе в большинстве случаев используется 20 аминокислот. Множество их комбинаций создают молекулы белков с большим разнообразием свойств.
Функции белков в клетках живых организмов более разнообразны, чем функции других биополимеров — полисахаридов и ДНК. Так, белки-ферменты катализируют протекание биохимических реакций и играют важную роль в обмене веществ. Некоторые белки выполняют структурную или механическую функцию, образуя цитоскелет, поддерживающий форму клеток. Также белки играют ключевую роль в сигнальных системах клеток, при иммунном ответе и в клеточном цикле.
Белки — важная часть питания животных и человека (основные источники: мясо, птица, рыба, молоко, орехи, бобовые, зерновые; в меньшей степени: овощи, фрукты, ягоды и грибы), поскольку в их организмах не могут синтезироваться все необходимые аминокислоты и часть должна поступать с белковой пищей. В процессе пищеварения ферменты разрушают потреблённые белки до аминокислот, которые используются для биосинтеза собственных белков организма или подвергаются дальнейшему распаду для получения энергии.
Определение аминокислотной последовательности первого белка — инсулина — методом секвенирования белков принесло Фредерику Сенгеру Нобелевскую премию по химии в 1958 году. Первые трёхмерные структуры белков гемоглобина и миоглобина были получены методом дифракции рентгеновских лучей, соответственно, Максом Перуцем и Джоном Кендрю в конце 1950-х годов, за что в 1962 году они получили Нобелевскую премию по химии.
Белки были выделены в отдельный класс биологических молекул в XVIII веке в результате работ французского химика Антуана де Фуркруа и других учёных, в которых было отмечено свойство белков коагулировать (денатурировать) под воздействием нагревания или кислот. В то время были исследованы такие белки, как альбумин («яичный белок»), фибрин (белок из крови) и глютен из зерна пшеницы.
В начале XIX века уже были получены некоторые сведения об элементарном составе белков, было известно, что при гидролизе белков образуются аминокислоты. Некоторые из этих аминокислот (например, глицин и лейцин) уже были охарактеризованы. Голландский химик Геррит Мульдер на основе анализа химического состава белков выдвинул гипотезу, что практически все белки имеют сходную эмпирическую формулу. В 1836 году Мульдер предложил первую модель химического строения белков. Основываясь на теории радикалов, он после нескольких уточнений пришёл к выводу, что минимальная структурная единица белка обладает следующим составом: C40H62N10O12. Эту единицу он назвал «протеином» (Pr) (от греч. протос — первый, первичный), а теорию — «теорией протеина». Сам термин «протеин» был предложен ещё шведским химиком Якобом Берцелиусом. Согласно представлениям Мульдера, каждый белок состоит из нескольких протеинных единиц, серы и фосфора. Например, он предложил записывать формулу фибрина как 10PrSP. Мульдер также исследовал продукты разрушения белков — аминокислоты и для одной из них (лейцина) с малой долей погрешности определил молекулярную массу — 131 дальтон. По мере накопления новых данных о белках теория протеина стала подвергаться критике, но, несмотря на это, до конца 1850-х всё ещё считалась общепризнанной.
К концу XIX века было исследовано большинство аминокислот, которые входят в состав белков. В конце 1880-х гг. русский учёный А. Я. Данилевский отметил существование пептидных групп (CO—NH) в молекуле белка. В 1894 году немецкий физиолог Альбрехт Коссель выдвинул теорию, согласно которой именно аминокислоты являются основными структурными элементами белков. В начале XX века немецкий химик Эмиль Фишер экспериментально доказал, что белки состоят из аминокислотных остатков, соединённых пептидными связями. Он же осуществил первый анализ аминокислотной последовательности белка и объяснил явление протеолиза.
Однако центральная роль белков в организмах не была признана до 1926 года, когда американский химик Джеймс Самнер (впоследствии — лауреат Нобелевской премии по химии) показал, что фермент уреаза является белком.
Сложность выделения чистых белков затрудняла их изучение. Поэтому первые исследования проводились с использованием тех полипептидов, которые легко могли быть очищены в большом количестве, то есть белков крови, куриных яиц, различных токсинов, а также пищеварительных/метаболических ферментов, выделяемых после забоя скота. В конце 1950-х годов компания Armour Hot Dog Co. смогла очистить килограмм бычьей панкреатической рибонуклеазы А, которая стала экспериментальным объектом для многих исследований.
Идея о том, что вторичная структура белков — результат образования водородных связей между аминокислотными остатками, была высказана Уильямом Астбери в 1933 году, но Лайнус Полинг считается первым учёным, который смог успешно предсказать вторичную структуру белков. Позднее Уолтер Каузман, опираясь на работы Кая Линдерстрём-Ланга, внёс весомый вклад в понимание законов образования третичной структуры белков и роли в этом процессе гидрофобных взаимодействий. В конце 1940-х — начале 1950-х годов Фредерик Сенгер разработал метод секвенирования белков, с помощью которого он к 1955 году определил аминокислотную последовательность двух цепей инсулина, продемонстрировав, что белки — это линейные полимеры аминокислот, а не разветвлённые (как у некоторых сахаров) цепи, коллоиды или циклолы. Первым белком, аминокислотную последовательность которого установили советские/российские учёные, стала в 1972 году аспартатаминотрансфераза.
Первые пространственные структуры белков, полученные методом дифракции рентгеновских лучей (рентгеноструктурного анализа) стали известны в конце 1950-х — начале 1960-х годов, а структуры открытые с помощью ядерного магнитного резонанса — в 1980-х годах. В 2012 году Банк данных о белках (Protein Data Bank) содержал около 87 000 структур белков.
В XXI веке исследование белков перешло на качественно новый уровень, когда исследуются не только индивидуальные очищенные белки, но и одновременное изменение количества и посттрансляционных модификаций большого числа белков отдельных клеток, тканей или целых организмов. Эта область биохимии называется протеомикой. С помощью методов биоинформатики стало возможно не только обработать данные рентгеноструктурного анализа, но и предсказать структуру белка, основываясь на его аминокислотной последовательности. В настоящее время криоэлектронная микроскопия крупных белковых комплексов и предсказание пространственных структур белковых доменов с помощью компьютерных программ приближаются к атомарной точности.
Свойства
Размер белка может измеряться в числе аминокислотных остатков или в дальтонах (молекулярная масса), но из-за относительно большой величины молекулы масса белка выражается в производных единицах — килодальтонах (кДа).
Белки обладают свойством амфотерности, то есть в зависимости от условий проявляют как кислотные, так и осно́вные свойства. В белках присутствуют несколько типов химических группировок, способных к ионизации в водном растворе: карбоксильные остатки боковых цепей кислых аминокислот (аспарагиновая и глутаминовая кислоты) и азотсодержащие группы боковых цепей основных аминокислот (в первую очередь, ε-аминогруппа лизина и амидиновый остаток CNH(NH2) аргинина, в несколько меньшей степени — имидазольный остаток гистидина). Каждый белок характеризуется изоэлектрической точкой (pI) — кислотностью среды (pH), при которой суммарный электрический заряд молекул данного белка равен нулю и, соответственно, они не перемещаются в электрическом поле (например, при электрофорезе).
Растворимость
Белки различаются по степени растворимости в воде. Водорастворимые белки называются альбуминами, к ним относятся белки крови и молока. К нерастворимым, или склеропротеинам, относятся, например, кератин (белок, из которого состоят волосы, шерсть млекопитающих, перья птиц и т. п.) и фиброин, который входит в состав шёлка и паутины. Растворимость белка определяется не только его структурой, но внешними факторами, такими как природа растворителя, ионная сила и pH раствора.
Денатурацией белка называют любые изменения в его биологической активности и/или физико-химических свойствах, связанные с потерей четвертичной, третичной или вторичной структуры (см. раздел «Структура белка»). Как правило, белки достаточно стабильны в тех условиях (температура, pH и др.), в которых они в норме функционируют в организме. Резкое изменение этих условий приводит к денатурации белка. В зависимости от природы денатурирующего агента выделяют механическую (сильное перемешивание или встряхивание), физическую (нагревание, охлаждение, облучение, обработка ультразвуком) и химическую (кислоты и щёлочи, поверхностно-активные вещества, мочевина) денатурацию.
Денатурация белка может быть полной или частичной, обратимой или необратимой. Самый известный случай необратимой денатурации белка в быту — это приготовление куриного яйца, когда под воздействием высокой температуры растворимый в воде прозрачный белок овальбумин становится плотным, нерастворимым и непрозрачным. Денатурация в некоторых случаях обратима, как в случае осаждения водорастворимых белков с помощью солей аммония, и используется как способ их очистки.
Структура
Молекулы белков представляют собой линейные полимеры, состоящие из остатков α-L-аминокислот (которые являются мономерами), также в состав белков могут входить модифицированные аминокислотные остатки и компоненты неаминокислотной природы. Для обозначения аминокислот в научной литературе используются одно- или трёхбуквенные сокращения. Хотя на первый взгляд может показаться, что использование в большинстве белков «всего» 20 видов аминокислот ограничивает разнообразие белковых структур, на самом деле количество вариантов трудно переоценить: для цепочки из 5 аминокислотных остатков оно составляет уже более 3 миллионов, а цепочка из 100 аминокислотных остатков (небольшой белок) может быть представлена более чем в 10130 вариантах. Белки длиной от 2 до нескольких десятков аминокислотных остатков часто называют пептидами, при большей степени полимеризации — белками, хотя это деление весьма условно.
При образовании белка в результате взаимодействия α-карбоксильной группы (-COOH) одной аминокислоты с α-аминогруппой (-NH2) другой аминокислоты образуются пептидные связи. Концы белка называют N- и C-концом, в зависимости от того, какая из групп концевого аминокислотного остатка свободна: -NH2 или -COOH, соответственно. При синтезе белка на рибосоме первым (N-концевым) аминокислотным остатком обычно является остаток метионина, а последующие остатки присоединяются к C-концу предыдущего.
Уровни организации
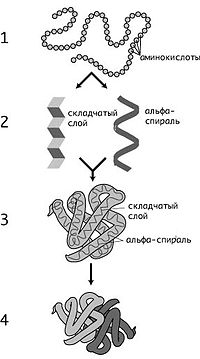
1 — первичная, 2 — вторичная, 3 — третичная, 4 — четвертичная
К. Линдстрём-Ланг предложил выделять 4 уровня структурной организации белков: первичную, вторичную, третичную и четвертичную структуры. Хотя такое деление несколько устарело, им продолжают пользоваться. Первичная структура (последовательность аминокислотных остатков) полипептида определяется структурой его гена и генетическим кодом, а структуры более высоких порядков формируются в процессе сворачивания белка. Хотя пространственная структура белка в целом определяется его аминокислотной последовательностью, она является довольно лабильной и может зависеть от внешних условий, поэтому более правильно говорить о предпочтительной или наиболее энергетически выгодной конформации белка.
Дата добавления: 2016-07-11; просмотров: 2692;
Поиск по сайту
Узнать еще
- БОЯЗНЬ АГРЕССИИ И ЗАЩИТНАЯ АГРЕССИЯ
- Внешняя молниезащитная система
- Воспаление как общая защитная реакция организма.
- Высоковольтная защитная и коммутационная аппаратура.
- Защитная и опознавательная окраска трубопроводов
- ЗАЩИТНАЯ ОДЕЖДА СПЕЦИАЛЬНАЯ ЗАЩИТНАЯ ОДЕЖДА (СЗО) СПАСАТЕЛЕЙ
- Защитная функция крови. Иммунитет. Регуляция иммунного ответа

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
