Динамика экосистем: сукцессии, флуктуации и климаксные сообщества
Любая экосистема постоянно изменяется и приспосабливается к колебаниям внешней и внутренней среды. Эта динамика может затрагивать отдельные компоненты, такие как популяции или трофические группы, либо всю систему в целом. Изменения бывают вызваны как внешними факторами, так и процессами, которые сама экосистема порождает и модифицирует. Таким образом, динамика является фундаментальным свойством существования любой экологической системы.
Наиболее простой тип динамики — суточный, связанный с ритмами фотосинтеза и транспирации у растений. Еще более выражены суточные изменения в поведении животного населения: разные виды проявляют активность в различное время суток. Аналогичные закономерности наблюдаются и в сезонных явлениях, которые еще сильнее влияют на активность и жизненные циклы организмов. Эти циклические изменения являются неотъемлемой частью функционирования биоценозов.
Многолетняя динамика также характерна для экосистем. На примере леса или луга легко заметить, что каждому году свойственны свои особенности — так называемые «клеверные» или «злаковые» годы. Это свидетельствует об индивидуальных требованиях каждого вида к условиям среды, где изменения могут быть благоприятны для одних и угнетать других. На эту динамику также накладывается периодичность в интенсивности размножения видов.
Эти многолетние изменения могут либо повторяться, нося циклический характер, либо быть однонаправленными, поступательными. Периодически повторяющуюся динамику называют флуктуациями (циклическими изменениями). Направленную динамику именуют поступательной, или развитием экосистем, что в конечном счете приводит к смене биоценозов. Этот процесс называется сукцессией (от лат. successio — преемственность).
Первичные сукцессии начинаются на изначально безжизненном субстрате, таком как песчаные дюны, скалы или застывшая лава. Общей закономерностью является постепенное заселение территории пионерными видами, увеличение биологического разнообразия и обогащение почвы органическим веществом. Происходит усложнение структуры биоценоза, уменьшение числа свободных экологических ниш и формирование все более устойчивых и продуктивных экосистем.
Каждая стадия сукцессионного ряда характеризуется своим набором видов, адаптированных к конкретным условиям. Последовательные стадии сменяют друг друга до формирования относительно устойчивого завершающего сообщества. Такое конечное состояние называют климаксным сообществом (от греч. klimax — лестница) или коренным биоценозом. Климаксные экосистемы отличаются стабильностью и способностью к самоподдержанию.
Американский эколог Фредерик Клементс, разработавший учение о сукцессиях, считал, что в рамках крупной природной зоны все сукцессионные ряды завершаются одним типом климакса (моноклимакс). Такой климакс определяется климатом региона и называется климатическим. Видовой состав этих экосистем может различаться, но их объединяют сходные виды-эдификаторы — доминантные виды, которые в наибольшей степени создают среду обитания.
Для степной зоны эдификаторами являются плотнокустовые злаки, например, ковыль и типчак. В лесной зоне Северной Евразии ключевыми эдификаторами выступают ель или пихта, которые сильно затеняют подпологовое пространство и подкисляют почву. Эти виды определяют условия для всего сообщества. С ними могут сосуществовать другие виды, не уступающие в скорости роста, что приводит к формированию смешанных лесов.
Перед формированием климакса экосистема проходит через ряд промежуточных стадий. В лесной зоне (Рис. 5) первыми на безжизненном субстрате появляются организмы-пионеры: накипные лишайники и водоросли. Их сменяют травянистые сообщества, затем светолюбивые кустарники и деревья-пионеры (береза, осина). Под их пологом создаются условия для поселения теневыносливых видов ели, которая со временем выходит в первый ярус, формируя климаксное сообщество.
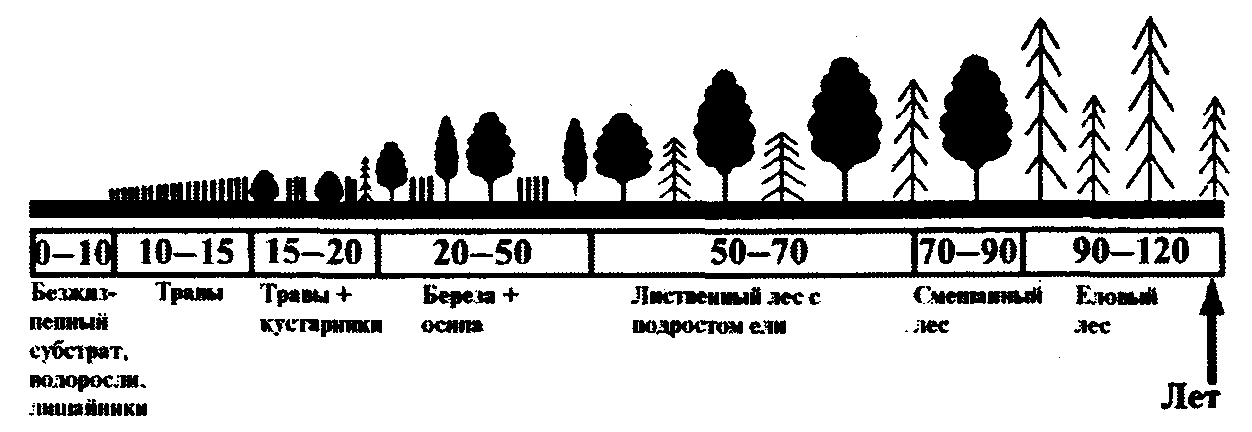
Рис. 5. Биогеоценотическая сукцессия на примере смен фитоценозов в лесной зоне
Наряду с теорией моноклимакса существует концепция поликлимакса, предполагающая, что в одном регионе может существовать несколько устойчивых завершающих сообществ. Например, в лесной зоне это могут быть луга или сосновые леса. Однако сторонники моноклимакса утверждают, что луга существуют лишь благодаря антропогенному воздействию (скашиванию), а сосняки — из-за бедности почв, не пригодных для ели. Со временем, по их мнению, эти сообщества все равно сменятся климатическим климаксом.
Причины сукцессий разнообразны. Иногда считают, что существующее сообщество создает неблагоприятные условия для себя самого (самоотравление, почвоутомление). Однако часто смена происходит без этого, как в случае внедрения ели под полог лиственных пород, где условия лишь улучшаются. Это объясняется тем, что ель использует ослабленное затенение под пологом березы или осины для своего развития.
В современном мире мощнейшим фактором динамики экосистем стал человек. Большинство коренных климаксных сообществ (например, степи) разрушены. Их место заняли производные, вторичные экосистемы. К сукцессионным сменам также ведут осушение болот, химическое загрязнение, рекреационные нагрузки, пожары и усиленный выпас скота. Эти воздействия часто упрощают структуру экосистем.
Антропогенные воздействия часто ведут к дигрессии — упрощению и деградации экосистем. Различают пастбищные, рекреационные и другие типы дигрессий. В отличие от сукцессии, дигрессия заканчивается не климаксом, а катоценозом — стадией глубокого угнетения, которая может привести к полному распаду системы. Климаксные экосистемы, такие как дубравы или хвойные леса, особенно чувствительны к таким вмешательствам.
Вторичные сукцессии отличаются от первичных тем, что начинаются не на безжизненном субстрате, а на месте нарушенных или разрушенных экосистем (после вырубок, пожаров, на заброшенных полях). Их главное отличие — значительно более высокая скорость протекания, так как они стартуют с промежуточных стадий и на богатых почвах. Однако если антропогенное давление продолжается, процесс сменяется дигрессией.
Выделяют также автотрофные и гетеротрофные сукцессии. Автотрофные (рассмотренные выше) протекают при участии продуцентов (растений) и потенциально бессмертны, так как подпитываются энергией солнца. Гетеротрофные сукцессии происходят в среде без продуцентов (например, при разложении трупа или бревна) и вовлекают только консументов и редуцентов. Они заканчиваются после полного исчерпания органического вещества.
Для любой сукцессии характерны общие закономерности. Вначале видовое разнообразие, биомасса и продуктивность невелики, но затем растут. Усиливаются взаимосвязи между организмами, усложняются трофические сети, сокращается число свободных экологических ниш. Интенсифицируются круговорот веществ и поток энергии. Скорость процесса зависит от продолжительности жизни организмов-эдификаторов.
В зрелой климаксной стадии биомасса достигает максимума, а продуктивность становится устойчивой. Существует мнение, что продуктивность климакса ниже, чем на промежуточных стадиях, из-за больших затрат на дыхание и потребления консументами. Однако для многих экосистем, особенно лесных, это не так — их продуктивность максимальна именно в климаксе из-за полного освоения пространства, хотя возможность изъятия продукции человеком может быть ограничена.
Сведения об авторах и источниках:
Автор: Н. А. Воронков
Источник: Основы общей экологии
Публикации предназначены для студентов всех специальностей, изучающих экологию как общекультурную дисциплину, а также для учителей, учащихся старших классов и всех интересующихся вопросами экологии.
Дата добавления: 2025-09-11; просмотров: 298;
Поиск по сайту
Узнать еще
- Гидродинамика в биологии и в простейшей медицинской аппаратуре
- Глобальное распределение атмосферного давления и циркуляция ветров: сезонная динамика и климатическое влияние
- Динамика численности популяций: рост, регуляция и гомеостаз
- Продуктивность экосистем: первичная и вторичная продукция, биомасса и их роль в биосфере
- Структура экосистем: от биогеоценоза до трофических цепей
- Устойчивость и стабильность экосистем: определение, различия и примеры

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
