ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ЦВЕТКА
В связи с тем, что цветковые растения играют большую роль в жизни человека, вопрос об их происхождении, а также о происхождении их специализированного репродуктивного органа – цветка – занимали и занимают в научных исследованиях ботаников разных профилей первостепенное место. Несмотря на то, что морфологи, систематики, эмбриологи, палеоботаники накопили большое количество фактического материала, до сих пор не потеряли своего значения слова великого натуралиста Ч. Дарвина «об ужасной тайне, окружающей цветковые растения».
Действительно, вопрос о морфологической природе частей цветка не разрешил проблемы происхождения цветка как особого репродуктивного органа.
Попытка найти возможных предков покрытосеменных растений и на основании изучения их репродуктивных органов попытаться найти репродуктивный орган, который мог бы быть предшественником цветка, «заставили» ботаников обратить внимание на ископаемые голосеменные. Так возникли псевдантовая (от греч. pseudo – ложь, anthos – цветок ) и эуантовая, или стробилярная (от греч. eu – истина, anthos – цветок) теории происхождения цветка.
Псевдантовая теория происхождения цветка была сформулирована Р. Веттштейном в1901 г. В качестве предковой группы покрытосеменных растений он рассматривал ископаемые голосеменные эфедровые, у которых типичных шишек нет, но существуют своеобразные структуры, которые называют «микростробилами» и «мегастробилами». Отдельный микростробил внешне похож на тычинку покрытосеменных, а группа микростробилов, окруженных кроющими листьями, -- на их соцветие. Мегастробил состоит из одного семязачатка (семяпочки) и окружен покровом. Р. Веттштейн полагал, что из микростробила в результате редукции кроющих листьев мог возникнуть тычиночный цветок с простым околоцветником, а из мегастробила – голый женский цветок. Согласно представлению Веттштейна, обоеполый цветок покрытосеменных гомологичен «соцветию» (собранию микро- и мегастробилов) эфедровых, но показать, каким образом из однополых цветков могли образоваться обоеполые, убедительно доказать не удалось. Постепенно ботаники от этой гипотезы отказались. Однако в свое время псевдантовая гипотеза сыграла определенную роль и была положена в основу ряда систем покрытосеменных растений. Самой известной из таких систем является система А. Энглера (1844 -- 1930), единственная система, филогенетические связи в которой прослеживаются до рода. До настоящего времени ею пользуются в ряде крупных Гербариев, в том числе и в Гербарии Ботанического института имени В. Л. Комарова Российской Академии наук (Санкт- Петербург).
К концу ХIХ началу ХХ веков создались предпосылки для осмысливания путей происхождения цветка в процессе эволюции. Опираясь на данные палеоботаники и морфологии, американский ботаник и филогенетик Ч. Бэсси (1845 - 1915) отмечает, что репродуктивный орган покрытосеменных состоит из стебля, на котором развиваются спороносные и стерильные листья, а позднее высказывает предположение, что покрытосеменные произошли от стробилоносных саговниковых. Независимо от Ч. Бэсси немецкий систематик Г. Галлир в 1902 г. на основании изучения позеленевшего пролиферирующего цветка водосбора (Aquilegia vulgaris) рассматривает цветок как видоизмененный репродуктивный побег. Таким образом, Ч. Бэсси и Г. Галлир фактически явились основоположниками эуантовой, или стробилярной, гипотезы происхождения цветка. Однако окончательно она была сформулирована несколько позже.
В 1906 г. Виланд описал обоеполый стробил ископаемых голосеменных – беннетитовых. Это открытие позволило Э. Арберу и Дж. Паркину окончательно сформулировать стробилярную гипотезу происхождения цветка. Согласно этой гтпотезе, цветок представляет собой обоеполый стробил, приспособленный для размножения. Цветок в представлении Э. Арбера и Дж. Паркина – это новейшая модификация стробила – антостробил. Э. Арбер Дж. Паркин считают, что беннетитовые, очевидно, не прямые предки покрытосеменных растений, но, возможно, они являются предками каких-то гипотетических полупокрытосеменных, которые до сих пор не выявлены. В своем труде «О происхождении покрытосеменных» (1907) авторы устанавливают признаки примитивности и специализации цветка и высказывают свою точку зрения на филогенетическую систему покрытосеменных. Она диаметрально противоположна точке зрения Р. Веттштейна. Если Р. Веттштейн считал наиболее примитивными семействами те, представители которых имели мелкие раздельнополые ветроопыляемые цветки без околоцветника или с редуцированным околоцветником, то, по Арберу и Паркину, самые примитивные семейства включают представителей с крупными обоеполыми насекомоопыляемыми цветками, имеющими простой или двойной околоцветник.
Открытие псилофитовых (риниообразных), признанных наиболее примитивными высшими растениями, дало основание вновь вернуться к проблеме происхождения цветка и выдвинуть теломную теорию. Основоположником ее является В. Циммерман (1930). Он считает, что все органы высших растений развились из простых и недифференцированных элементов, названных им теломами. Согласно представлениям В. Циммермана, стерильные телломы – филлоиды и фертильные теломы, несущие спорангии, могли срастаться и формировать синтеломы. Цветок, по В. Циммерману, представляет собой синтелом, процесс возникновения которого происходил параллельно процессу формирования вегетативного побега. И хотя детально эта теория не разработана, она имела большое значение для решения некоторых вопросов о природе элементов цветка. С 40-х – 50-х годов ХХ столетия – периода особенно острых дискуссий сторонников теломной теории и приверженцев классической морфологии, цветок перестали определять как «метаморфизированный вегетативный побег».
В связи с появлением новых научных данных появляются и новые гипотезы происхождения цветка, однако, фактически ни одна из них не является достаточно убедительной. Суммируя все знания в этой области известный советский ботаник Б. М. Козо-Полянский (1890 -1957) вынужден был констатировать: «Не известны ни предок цветковых растений, ни конкретный орган, исторически исходный для цветка».
ОБЩИЙ ПЛАН СТРОЕНИЯ ЦВЕТКА
Цветки покрытосеменных растений чрезвычайно разнообразны по строению, форме, величине, однако у них имеются четко выраженные общие признаки. В состав цветка входит цветоложе, околоцветник (иногда он отсутствует), тычинки и пестики или один из этих элементов. Очень редко тычинки и пестики в цветке отсутствуют (воронковидные цветки василька – Centaurea cyanus).
Цветоложе (лат. receptaculum) – осевая часть цветка, на которой располагаются все его элементы. Цветоложе может быть вытянутым – коническим, что подтверждает его осевое происхождение, или укороченным – плоским. Коническое цветоложе встречается у представителей примитивных семейств (сем. Magnoliaceae – Магнолиевые, Ranunculacea – Лютиковые). У большинства же покрытосеменных растений цветоложе плоское, а у представителей семейства Розоцветные (Rosaceae) оно блюдцевидно расширенное (Fragaria vesca – Земляника лесная) или бокальчато вогнутое (Prunus vulgaris – Слива обыкновенная) и называется гипантий.
Элементы цветка расположены на цветоложе в определенном порядке. Все элементы цветка можно подразделить на стерильные (не принимающие непосредственного участия в репродуктивных процессах) и фертильные (в которых происходят репродуктивные процессы). К стерильным элементам относится околоцветник цветка, к фертильным – тычинки и пестики.
В зависимости от строения околоцветник цветка, или его покровы (лат. Periantium), может быть двойным и простым. Двойной околоцветник состоит из чашечки (лат. Calyx) и венчика (лат. Corolla). Чашечка образована чашелистиками. Чашелистики имеют листовую природу, о чем свидетельствуют анатомические данные. Для чашелистиков характерно наличие трех листовых следов, т.е. такое же число листовых следов, которое типично и для вегетативных листьев. Чашелистики могут быть не сросшимися, тогда образуется свободнолистная чашечка (Ranunculus acris – Лютик едкий, Stellaria graminea – Звездчатка злаковидная). Если чашелистики в той или иной степени срастаются, формируется сростнолистная чашечка (Agrostemma githago – Куколь обыкновенный,
Veronica chamaedrys – Вероника дубравная). Чашелистики чаще всего окрашены в зеленый цвет, но могут иметь и иную окраску: желтую (Лютик едкий), синюю (Delphinium grandiflorum – Живокость крупноцветковая), оранжевую (Trollius asiaticus – Купальница азиатская), белую, розовую, фиолетовую (Hydrangea hortensis – Гортензия садовая). Чашелистики на стадии бутона выполняют защитную функцию, а после раскрытия бутона зеленые чашелистики принимают участие в процессе фотосинтеза, а ярко окрашенные – в привлечении насекомых-опылителей.
За чашечкой располагается венчик, состоящий из лепестков. Как и чашечка, он может быть свободнолепестным (Malus domestica – Яблоня домашняя, Sedum acre – Очиток едкий) и спайнолепестным (Campanula media – Колокольчик средний, Digitalis grandiflora – Наперстянка крупноцветковая). Морфологическая природа лепестков различными авторами трактуется по-разному. Одни авторы (Гёте и его последователи) считают, что лепестки произошли, как и чашелистики, из верхушечных листьев, другие (Декандоль и др.) считают их производными тычинок. Э. Арбер, Дж. Паркин, Б. М. Козо-Полянский, А. Имс полагают, что лепестки могут иметь двоякое происхождение. Подтверждением тому являются данные васкулярной анатомии. Так у некоторых лютиковых лепестки, как и тычинки, имеют только один листовой след, в то время как у представителей семейств магнолиевых (сем. Magnoliaceae), лимонниковых (сем. Schisandraceae) лепестки, как и чашелистики, имеют по три листовых следа. Лепестки выполняют защитную функцию, а также функцию привлечения насекомых-опылителей. Привлечению насекомых-опылителей способствует не только яркая окраска лепестков венчика, но и образование на лепестках (виды рода Batrachium – Шелковник) или из лепестков (роды Aconitum – Борец, Aquilegia – Водосбор) различных по строению нектарников.
Если околоцветник однородно окрашен, его называют простым. В зависимости от окраски различают чашечковидный (Beta vulgaris – Свекла обыкновенная, Chenopodium album – Марь обыкновенная) и венчиковидный (Narcissus poeticus – Нарцисс поэтический, Colchicum autumnale – Безвременник осенний) околоцветник. Однако иногда простой околоцветник, образованный чашелистиками, бывает венчиковидно окрашенным (Aconitum napellus – Борец аптечный, Consolida regalis – Сокирки обыкновенные).
Важнейшими элементами цветка являются тычинки и пестики. Совокупность тычинок составляет андроцей (Androeceum) цветка. В зависимости от строения различают андроцей однобратственный, двубратственный и многобратственный. В однобратственном андроцее все тычинки сращены в одну группу (Malva excisa – Мальва разрезнолистная, Lupinus polyphyllus – Люпин многолетний). Если тычинки срастаются в две группы, андроцей называется двубратственным (Vicia cracca –Горошек мышиный, Cucurbita pepo – Тыква обыкновенная). Если формируется неколько групп сросшихся тычинок (Hypericum perforatum – Зверобой продырявленный) или если все тычинки свободные (Pulsatilla patens – Прострел раскрытый), андроцей считается многобратственным. Андроцей может быть двусильным, тогда в цветке из четырех тычинок 2 длинные, а 2 короткие (сем. Labiatae – Губоцветные) или четырехсильным – в цветке 4 тычинки длинные, а 2 короткие (сем. Cruciferae – Крестоцветные).
Тычинка состоит из тычиночной нити, пыльника, состоящего из двух половинок (двух тек), и связника – паренхимной ткани, которая соединяет половинки пыльника. Тычинка представляет собой редуцированный микроспорофилл, остатком которого являются тычиночная нить и связник, с микроспорангиями, находящимися в пыльнике.
Сравнительное изучение тычинок различных представителей покрытосеменных позволило составить представление об эволюции этого репродуктивного элемента. У современных примитивных покрытосеменных (особенно у рода Дегенерия, некоторых магнолий) тычинки устроены очень архаично: они плоские, широкие, не расчлененные на тычиночную нить, пыльник и связник с линейными микроспорангиями, расположенными ламинально, т.е. на поверхности пластинчатого микроспорофилла. Пыльники располагаются как на адаксиальной (внутренней) стороне и тогда вскрываются интрозно, так и на абаксиальной стороне. В этом случае они вскрываются экстрозно. В примитивных микроспорофиллах имеется три хорошо развитых проводящих пучка (три жилки). Постепенно в процессе эволюции микроспорофилл редуцировался, уменьшалось число проводящих пучков в мезофилле, исчезла большая часть мезофилла и в конечном счете сформировалась специализированная тычинка, дифференцированная на тычиночную нить, пыльник и связник. В тычиночной нити проходит 1 проводящий пучок, который доходит до связника и поставляет к живым паренхимным клеткам связника питательные вещества. По мере дифференцировки тычинки меняется положение пыльника. Постепенно половинки пыльника перемещаются в боковое положение, а затем вследствие дальнейшей редукции связника сливаются в пыльник, состоящий из двух половинок (двух тек), который занимает верхушечное положение. Таким образом, у большинства покрытосеменных пыльник содержит 4 микроспорангия и представляет собой синангий (совокупность спорангиев).
У некоторых растений в результате приспособления к опылению часть тычинок утрачивает пыльники и превращается в нектарники – стаминодии (Parnassia palustris – Белозор болотный, Trollius europaeus – Купальница европейская).
Число тычинок в обоеполых цветках варьирует в широких пределах – от бесконечно большого (сем. Ranunculaceae –Лютиковые, Magnoliaceae -- Магнолиевые) до двух (Veronica chamaedrys – Вероника дубравная) и даже одной (Platanthera bifolia – Любка двулистная). В однополых – тычиночных цветках количество тычинок также бывает различным.
Тычинки в цветке располагаются или по спирали (сем. Магнолиевые, большинство представителей сем. Лютиковые), или по кругу (сем. Rosaceae – Розоцветные, Caryophyllaceae -- Гвоздичные).
Центральную часть цветка занимает один или несколько пестиков. Совокупность пестиков составляет гинецей (Gynoeceum) цветка. Пестик представляет собой видоизмененный и высоко специализированный мегаспорофилл (плодолистик, карпелла) с мегаспорангиями (семяпочками, семязачатками), дифференцированный на завязь, стилодии (stylodium –суженная часть плодолистика) или столбик (stylos –образование, возникающее в результате срастания нескольких стилодиев) и рыльце.
Листовая природа плодолистика была научно обоснована О. П. Декандолем еще в 1827 г., хотя он ошибочно выводил их не из спороносных, а из вегетативных листьев. И действительно, чем примитивнее цветок, тем более листовидный характер носят отдельные плодолистики, и проводящая система их сходна с проводящей системой листа. Согласно представлениям А. Л. Тахтаджяна, от открытого развернутого мегаспорофилла плодолистик покрытосеменных отличается тем, что он как бы сложен по центральной жилке, т.е. сдвоенный (кондупликатный). В связи с кондупликатностью плодолистика мегаспорангии (семяпочки) оказываются на адаксиальной (внутренней) стороне, что повысило уровень их защищенности от различных неблагоприятных воздействий (сухости воздуха, высоких и низких температур, резких перепадов температур, повреждения насекомыми и т.д.).
Постепенно сближенные края кондупликатного плодолистика начинали срастаться, что привело в конечном счете к образованию типичного замкнутого пестика. Среди современных покрытосеменных самые архаичные пестики обнаружены у Дегенерии. Они срастаются только в основании. Верхние же края их не только не сращены, но фактически даже не соприкасаются друг с другом (Тахтаджян, 1964). Неполное срастание краев плодолистиков наблюдается также и у некоторых более продвинутых представителей (Paeonia – Пион, Platanus -- Платан). У резедовых завязь на верхушке остается незамкнутой.
Так как в кондупликатном плодолистике семяпочки оказались закрытыми и недоступными для пыльцы, возникла необходимость в образовании специальной поверхности, которая могла бы воспринимать пыльцу. Так постепенно у покрытосеменных на краях кондупликатных плодолистиков начала формироваться особая железистая ткань и возникла рыльцевая поверхность, или рыльцевый гребень. Наличие рыльцевого гребня – одна из характернейших особенностей мегаспорофиллов примитивных покрытосеменных. Сначала рыльцевый гребень простирался по обоим краям сложенного широкого плодолистика, но постепенно по мере срастания краев он укорачивался, менялась его форма, и в конечном счете он превратился в рыльце пестика, которое заняло верхушечное положение. В ходе эволюции дифференцировался и сам плодолистик, постепенно превратившийся в типичный пестик. Верхняя часть его вытянулась и превратилась в стилодий – стерильное образование. У примитивных покрытосеменных стилодий еще кондупликатный с двумя рыльцевыми гребнями, но постепенно края его срастаются, а на верхушке образуется типичное рыльце.
Пестик может быть образован одним или несколькими сросшимися плодолистиками. Гинецей, в котором пестик образован одним сросшимся плодолистиком, называется апокарпным. Апокарпный гинецей может состоять из нескольких или большого числа пестиков (сем. Магнолиевые, большинство лютиковых) или только одного пестика (сем. Fabaceae – Бобовые, Cerasus vulgaris – Вишня обыкновенная). Таким образом апокарпный гинецей может быть многочленным и одночленным. Многочленный апокарпный гинецей характерен для представителей примитивных семейств, одночленный встречается у эволюционно более продвинутых.
В процессе эволюции происходило усложнение строения гинецея. Благодаря срастанию нескольких плодолистиков сформировался ценокарпный гинецей. Число сросшихся плодолистиков в ценокарпном гинецее варьирует от двух (сем. Cruciferae – Крестоцветные, Gramineae -- Злаки) до неопределенно большого (род Malva – Мальва, или Просвирник). В зависимости от характера срастания плодолистиков ценокарпный гинецей подразделяется на синкарпный, паракарпный и лизикарпный. В синкарпном гинецее плодолистики полностью срастаются боковыми (латеральными) сторонами, в результате чего образуется многогнездная завязь, число гнезд в которой соответствует числу сросшихся плодолистиков (2 – у Hyoscyamus niger – Белены черной, 5 – у Linum usitatissimum – Льна посевного).
В паракарпном гинецее плодолистики срастаются таким образом, что завязь остается одногнездной (сем. Papaveraceae – Маковые, Cruciferae – Крестоцветные, Cucurbitaceae – Тыквенные). В лизикарпном гинецее завязь также одногнездная, но он возникает не так, как паракарпный. При образовании лизикарпного гинецея перегородки (стенки сросшихся плодолистиков) разрушаются, но в центре завязи остается колонка – центральная часть сросшихся плодолистиков (сем. Caryophyllaceae –Гвоздичные).
Эволюционное взаимоотношение различных типов гинецея можно представить следующим образом (рис. 1).
При развитии разных типов ценокарпного гинецея стилодии или остаются совершенно свободными (Сarum carvi – Тмин обыкновенный, Stellaria media – Звездчатка средняя) или частично срастаются (Malus domestica – Яблоня домашняя). У многих видов образуются столбики (Raphanus raphanistrum – Редька дикая, Veronica officinalis – Вероника лекарственная).
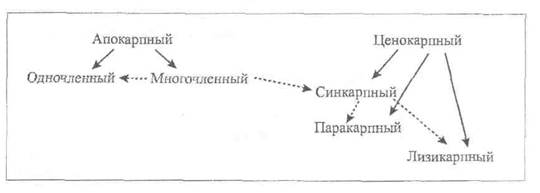
Рис. 1. Схема классификации (→) гинецея и предполагаемых эволюционных связей (----►) разных типов гинецея
В зависимости от типа гинецея типы плацентации (способы прикрепления) семяпочек различны. Все виды плацентации можно подразделить на ламинальную (поверхностную) и сутуральную (шовную). Самым примитивным типом плацентации является диффузная (ламинально-диффузная) плацентация, при которой семяпочки рассеяны по всей адаксиальной поверхности завязи (Butomus umbellatus – Сусак зонтичный). В апокарпном гинецее плацентация может быть медианной (ламинально-медианной) и латеральной (ламинально-латеральной). При медианной плацентации семяпочки располагаются вдоль средней жилки плодолистика (род Nelumbo – Лотос). При латеральной плацентации семяпочки располагаются вдоль брюшного шва, т.е. по краю сросшегося плодолистика (Pisum sativum – Горох посевной).
Сутуральные типы плацентации подразделяются на центрально-угловую, париэтальную (постенную) и центрально-осевую (колончатую). Центрально-угловая плацентация характерна для синкарпного гинецея. При таком типе плацентации семяпочки прикреплены в углах сросшихся плодолистиков (Lilium tigrinum – Лилия тигровая, Pyrus communis – Груша обыкновенная). В паракарпном гинецее плацентация париэтальная, но семяпочки могут прикрепляться по-разному. Если они располагаются по краям сросшихся плодолистиков, париэтальная плацентация называется латеральной (постенно-краевой), если вдоль центральной жилки – медианной (постенно-спинной) (Viola tricolor – Фиалка трехцветная, Cucumis sativus – Огурец посевной).
В лизикарпном гинецее семяпочки прикрепляются к колонке, и плацентация является центрально-осевой (роды Dianthus – Гвоздика, Saponaria – Мыльнянка).
В процессе развития гинецея плаценты могут сильно разрастаться и образовывать ложные перегородки (сем. Крестоцветные).
Число семяпочек как в апокарпном, так и в ценокарпном гинецее, варьирует от многих до одной. Уменьшение числа семяпочек в завязи пестика, как и уменьшение, а также усложнение строения пестика, – результат эволюционной специализации представителей различных семейств.
Как результат процесса эволюции можно рассматривать и различное положение завязи в цветке. Положение завязи определяют по отношению к околоцветнику цветка. Если околоцветник прикреплен под завязью, завязь называется верхней. Такая завязь только основанием прикреплена к цветоложу (роды Fragaria – Земляника, Ranunculus – Лютик, Secale – Рожь). Если околоцветник находится над завязью, завязь называется нижней (сем. Campanulaceae – Колокольчиковые, Umbelliferaе – Зонтичные, Iridaceae – Касатиковые). Завязь может быть полунижней, тогда околоцветник прикреплен в центральной части завязи (Lonicera tatarica – Жимолость татарская). У вишни обыкновенной (Cerasus vulgaris), черешни (Cerasus avium) свободная завязь глубоко «вдвинута» в цветоложе-гипантий. Такую завязь рассматривают как среднюю. Иногда понятия «полунижняя» и «средняя» завязь рассматривают как синонимы. Наиболее примитивным типом завязи считается верхняя завязь.
Дата добавления: 2022-07-20; просмотров: 199;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
