ПАЛЕОНТОЛОГИЧЕСКИЕ (БИОСТРАТИГРАФИЧЕСКИЕ) МЕТОДЫ
Более надежными методами определения относительного возраста отложений и корреляции последних на больших территориях являются палеонтологические методы, основанные на использовании ископаемых остатков организмов.
В основе палеонтологических методов лежит принцип последовательной смены неповторяющихся в разрезах фаунистических и флористических комплексов, отражающих этапность их исторического развития. Преимущество этих методов стратиграфии перед другими состоит в том, что они базируются на одном из основных положений эволюционной теории — неповторяемости эволюции. Необратимость эволюционного процесса впервые установил Ч. Дарвин. Он указал, что исчезнувший вид никогда не может появиться снова. Бельгийский ученый Л. Долло в 1893 г. положение о необратимости эволюции выдвинул в качестве «закона эволюции», согласно которому организм не может вернуться хотя бы частично к прежнему состоянию, которое было уже осуществлено в ряду его предков.
Исходя из закона необратимости эволюции Дарвина — Долло каждый комплекс ископаемых остатков организмов, приуроченный к тому или иному слою, отражает определенный этап развития органического мира и является неповторимым. Это и дает возможность использовать ископаемые остатки организмов для определения относительного возраста горных пород.
В основе палеонтологических методов лежит также явление широкого пространственного распространения ископаемых остатков организмов, что позволяет осуществлять корреляцию разрезов отдаленных друг от друга областей. Широкое пространственное распространение отдельных групп организмов связано с большой скоростью их расселения по поверхности Земли. Время, необходимое для расселения организмов, в геологических масштабах практически ничтожно при условии, если организмы не встречают на своем пути различного рода препятствий.
Определение возраста горных пород (их датировка) производится путем сравнения окаменелостей из этих пород с теми, которые встречаются в опорном разрезе. Например, в горной породе найден коралл Calceola sandalina, характерный для эйфельского яруса среднего девона в его стратотипическом (опорном) разрезе Западной Европы. Следовательно, и слой, в котором обнаружен коралл Calceola sandalina, относится также к эйфельскому ярусу. При расчленении и корреляции разрезов надо знать вертикальное распределение ископаемых остатков организмов по разрезу. Иногда они встречаются от подошвы до кровли и тогда полученные данные о возрасте будут относиться ко всему этому слою. Чаще же окаменелости встречаются спорадически, т. е. приурочены к определенным уровням в изучаемом слое, и возраст, установленный по таким формам, условно распространяется на весь слой.
Ценность для биостратиграфии отдельных групп ископаемых остатков организмов далеко не одинакова. Она определяется распространением этих групп во времени и в пространстве, степенью приуроченности к определенным типам пород, частотой встречаемости в горных породах и темпами их эволюции. В связи с этим выделяют архистратиграфические (ортостратиграфические) и парастратиграфические группы вымерших организмов. Архистратиграфические группы, характеризующиеся быстрой эволюцией, широким распространением и независимостью от осадка4 (пород), к которым относят, например, гониатиты, аммониты, граптолиты, планктонные фораминиферы и др., позволяют проводить не только детальное расчленение разреза, но и не менее детальную корреляцию с опорными разрезами. Это, как правило, нектонные и планктонные организмы, скелеты которых могут захороняться на любом участке морского дна, независимо от осадка. К парастратиграфичеоким группам относят, как правило, бентосные организмы, например брахиоподы, кораллы, мшанки и др., которые благодаря тесной связи с определенными типами осадков в разных районах образуют различные по составу ассоциации. Они в меньшей степени могут быть использованы для широких сопоставлений, но для определения возраста пород и корреляции в пределах районов распространения этих групп имеют иногда первостепенное значение.
Деление вымерших организмов на архистратиграфические и парастратиграфические группы в значительной мере условно и отражает степень наших знаний об этих группах. Так, например, до недавнего времени радиолярии считались второстепенной парастратиграфической группой, а в настоящее время, благодаря использованию электронной микроскопии, они стали одной из основных групп для определения возраста и корреляции верхнемеловых и кайнозойских отложений.
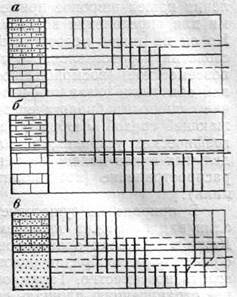
Биостратиграфическое расчленение частных разрезов производят путем анализа распространения в них отдельных таксонов вымерших организмов или их комплексов. Наиболее простой моделью является следующая: в каждом последовательно сменяющемся слое содержится свой вид — «а», «б», «в». По этим видам выделяют биостратиграфические подразделения Л, Б, В, границы которых совпадают с границами слоев или пачек (рис. 7,а). Однако чаще наблюдается несовпадение границ распространения отдельных видов или комплексов с границами слоев, что отражается и в несовпадении биостратиграфических и литологических границ (см. рис. 7,6). Какие же существуют критерии проведения границ биостратиграфических подразделений? Одни исследователи проводят их по появлению новых таксонов (вида, или группы видов), другие — по исчезновению старых, третьи считают границы, проведенные по появлению или исчезновению таксонов, равноценными (рис. 8,а). В геологической практике часто отдают предпочтение той границе, которая совпадает с литологической, хотя такая граница с биостратиграфических позиций может оказаться менее обоснованной (см. рис. 8,6), чем другие. Первостепенное значение приобретает та граница, которая проведена по появлению видов в одном филогенетическом ряду, т. е. видов, имеющих прямую родственную связь с видами подстилающих слоев (см. рис. 8,в).
Существует несколько биостратиграфических методов определения возраста горных пород. Ниже рассмотрены некоторые из них.
|
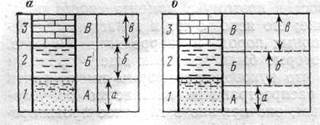 |
Рис. 7. Примеры распределения видов «а», «б», «s» в разрезах
/, 2, 3 — литологические подразделения; А, Б, В — биостратиграфические подразделения
Рис. 8. Различные варианты проведения границ биостратиграфических подразделений (справа за рамкой показаны предпочтительные уровни). По Д. Л. Степанову и М. С. Месежникову (1979 г.), с изменениями
Метод руководящих форм. Руководящими формами называют остатки вымерших животных и растений, которые отвечают определенным требованиям: они должны иметь небольшое вертикальное распространение по разрезу пород, но широкое в горизонтальном направлении (географическое), встречаться в слоях в достаточно большом количестве экземпляров, иметь хорошую сохранность и характерные особенности строения, позволяющие легко их определять. Обычно руководящие формы приурочены только к определенному слою, поэтому по ним легко определить относительный возраст слоя, в котором они находятся.
Этот метод является первым палеонтологическим методом, который был введен в стратиграфию. Он возник на рубеже XVIII и XIX веков и был впервые применен У. Смитом при расчленении и корреляции юрских отложений Англии. Научное обоснование этот метод получил в середине прошлого столетия, когда немецкий палеонтолог Г. Брони ввел понятие о руководящих формах и составил первый атлас руководящих форм беспозвоночных.
Можно назвать многие виды вымерших организмов (аммонитов, белемнитов, брахиопод, граптолитов, трилобитов, фораминифер), которые отвечают всем перечисленным выше требованиям. Так, например, аммонит Cardioceras cor datum является руководящим для оксфордского яруса верхней юры; белемнит Belemnitella mucronata — для кампанского яруса верхнего мела. Роды и более крупные систематические группы (семейства, отряды и даже типы) также могут быть руководящими для более крупных, чем ярус, стратиграфических подразделений — отделов, систем. К таким ископаемым относятся, например, археоциаты (Archaeocyathi), составляющие самостоятельный тип вымерших раннекембрийских организмов.
Сущность метода руководящих форм состоит в том, что из общего
количества окаменелостей, встреченных в определенном слое разреза,
выбирают один или 2—3 характерных для этого слоя видов, которые
отвечают указанным выше требованиям. Эти виды и будут руководящими формами для рассматриваемого слоя. Согласно этому методу
одновозрастными будут слои, содержащие идентичный или близкий
состав руководящих форм.
Рассматриваемый метод прост и поэтому с момента своего появления нашел широкое применение; его используют и в настоящее время для определения возраста горных пород и корреляции разрезов. Но он не лишен недостатков, которые были замечены еще в прошлом столетии. Представление о том, что руководящие формы имеют почти повсеместное (космополитное) распространение независимо от условий обитания, оказалось неверным, оно пришло в глубокое противоречие с данными биологической науки о тесной связи организма с окружающей средой. Любой организм обитает в определенных физико-географических условиях, поэтому пространственное распространение отдельных видов далеко не одинаково. Одни виды имеют очень широкое распространение (космополиты), другие — весьма ограниченное (эндемики).
В природе не существует абсолютных космополитов, так как ни один вид не может одновременно обитать на континентах и в морях. Даже в пределах одного континента или морского бассейна в силу большого разнообразия условий один и тот же вид не может существовать повсеместно. Таким образом, каждый вид имеет более или менее ограниченную площадь своего распространения (ареал).
Несостоятельным оказалось и второе положение, лежащее в основе метода руководящих форм,— приуроченность их во всех районах к одному и тому же стратиграфическому уровню. Это положение предусматривает повсеместное одновременное появление и вымирание руководящих форм. Как известно, один и тот же вид не может возникнуть сразу во многих районах земной поверхности. Он появляется на одном участке, а затем расселяется в другие районы. Однако скорость расселения организма зависит не только от его способности к перемещению, но и в значительной мере от возможности преодолевать различные препятствия, встреченные на путях миграции.
Препятствия могут быть биологические (например, отсутствие пищи) и физические. Для наземных животных и растений физическими препятствиями являются обширные пространства морей, неблагоприятный климат и непреодолимые элементы рельефа ^например, высокие горные хребты); для морских — участки суши (материки, острова), морские течения с неблагоприятной температурой, глубоководные впадины (для бентосных организмов) и т. д. Изменение физико-географической обстановки способствует заселению организмами новых районов, поэтому руководящий вид может занимать в разных районах различное стратиграфическое положение и в таком случае разновозрастные отложения с одинаковыми руководящими формами могут быть ошибочно приняты за одновозрастные.
Нельзя не учитывать также и другие явления, осложняющие применение метода руководящих форм. Одним из них является рекурренция, т. е. присутствие одних и тех же руководящих форм или целых комплексов в различных стратиграфических горизонтах. Явление рекурренции связано с миграцией организмов из района первоначального обитания в связи с изменившимися условиями в смежные районы и возвращением их через некоторый промежуток времени на старое место обитания, где к этому моменту восстановились благоприятные условия (рис. 9). Поэтому использование при определении геологического возраста и корреляции только руководящих форм без учета сопутствующего комплекса ископаемых остатков организмов может привести к грубым ошибкам в стратиграфических выводах. 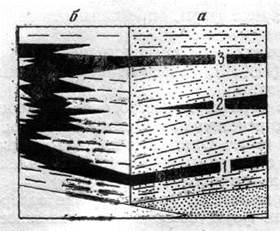
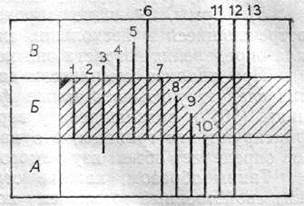 |
Рис. 10
Рис. 9. Схема, показывающая появление рекуррентных комплексов фауны в разновозрастных, но одинаковых по ли-тологическим признакам осадках. По Р. Муру (1948 г.)
а — комплекс фауны в черных сланцах, характерный для слоя 1, повторяется без существенных изменений выше по разрезу (слои 2 и 3); б — непрерывность накопления черных сланцев в условиях многократного перемещения зоны седиментации
Рис. 10. Пример использования комплекса форм одной группы вымерших организмов для расчленения разреза. Заштриховано изучаемое стратиграфическое подразделение
Метод анализа фаунистических и флористических комплексов. В отличие от метода руководящих форм при анализе фаунистических и флористических комплексов используют весь палеонтологический материал или определенные группы вымерших организмов. Одно из важных преимуществ этого метода состоит в том, что корреляция и выводы о геологическом возрасте базируются не на единичных, иногда случайно выбранных формах (видах), а на совокупности всех форм выбранной группы или различных групп организмов. Важно также и то, что данные о возрасте, полученные при анализе комплекса одной группы организмов, можно контролировать по другим группам; это снижает вероятность ошибки. Поэтому метод комплексов нашел широкое практическое применение и в настоящее время является основным при биостратиграфических исследованиях.
При анализе комплекса форм вымерших организмов могут быть выделены (рис. 10):
1) формы, встречающиеся только в данном слое и не переходящие
его границы (1, 2), эти формы являются руководящими; как правило,
они составляют небольшой процент в составе комплекса, но имеют
наибольшее стратиграфическое значение;
2) формы, появляющиеся в подстилающем слое и исчезающие в
вышележащем (3), но в данном слое встречающиеся часто и в боль
шом количестве; время существования этих форм длительное, но вре
мя их расцвета совпадает со временем формирования данного слоя;
такие формы называют характерными, или контролирующими;
3) формы, впервые появившиеся в данном слое и переходящие в
вышележащие слои (4, 5, 6); по таким формам обычно проводится
нижняя граница стратиграфического подразделения;
4) формы, заканчивающие свое существование во время форми
рования изучаемого слоя и обычно приуроченные к его нижней части
(7, 8, 9); такие формы называются доживающими; они имеют гораздо меньшее значение, чем предыдущие, кроме форм, определяющих верх
нюю границу (7);
5) формы, имеющие широкое вертикальное распространение, т. е. встречающиеся в нескольких слоях (11, 12), называют транзитными; для определения возраста они не пригодны.
При анализе комплексов данного стратиграфического подразделения следует учитывать также формы, исчезнувшие у его нежней границы (10) и появившиеся у верхней (13). Они, как правило, четко фиксируют эти границы, что наряду с другими признаками комплексов определяет объем изучаемого подразделения.
Таким образом, каждое стратиграфическое подразделение содержит свойственный только ему комплекс форм, позволяющий надежнее, чем по отдельным формам, определять геологический возраст и устанавливать границы этого подразделения. Каждый фаунистический или флористический комплекс неповторим в разрезе, так как эволюция органического мира необратима.
Следует помнить, что изменчивость комплекса форм во времени зависит от многих причин, главной из них является изменение. Внешних условий. Отдельные группы организмов, входящие в состав комплекса, по разному реагируют на эти изменения: одни из них меняются -быстро, другие — медленно. Поэтому рубежи этих изменений, а следовательно, и положение границ стратиграфических подразделений по разным группам вымерших организмов могут не совпадать.
Процентно-статистический метод был введен в стратиграфию Ч. Лайелем в прошлом столетии и широко применялся до 30-х годов нашего века. Ч. Лайель применил процентно-статистический метод для стратиграфического расчленения и корреляции кайнозойских отложений Западной Европы. На основании процентного содержания современных видов моллюсков в этих отложениях он выделил эоцен, миоцен и плиоцен. К эоцену были отнесены отложения, содержащие 3,5% ныне живущих видов моллюсков, к миоцену 17%, к нижнему плиоцену 35—50%, а к верхнему плиоцену 90—95%.
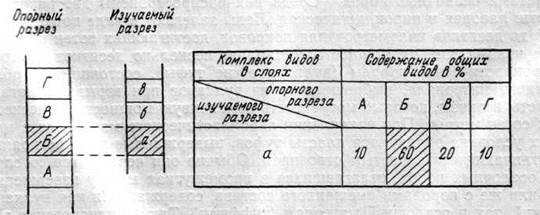
Процентно-статистический метод стали вскоре после его появления широко применять при расчленении и корреляции не только кайнозойских, но и более древних отложений. Определение относительного возраста того или иного слоя основывалось на подсчете процентного содержания общих видов с эталонным, детально изученным разрезом близких по возрасту отложений. Одновозрастными с эталонными считались те слои, которые содержали наибольший процент общих видов. Например, в эталонном разрезе (рис. 12) выделены четыре слоя А, Б, В, Г и для каждого слоя установлен определенный комплекс видов. В изученном разрезе выделены три слоя а, б, в. Из этих слоев также определен видовой состав вымерших организмов. Затем состав видов слоя «а» сравнивают последовательно с составом видов слоев А, Б, В, Г опорного разреза и подсчитывают процент общих видов. Пусть в слое «а» определено 50 видов, из которых б видов общих со слоем А (10%), 30 видов —со слоем Б (60%), 10 видов —со слоем В (20%) и 5 видов —со слоем Г (110%). По процентному содержанию общих видов слой «а» изучаемого разреза принимают одновозрастным со слоем Б опорного разреза. Такой же подсчет проводят для остальных слоев.
Следует отметить, что процентно-статистический метод является формальным, его применение не раз приводило к ошибкам в стратиграфии. По мере накопления фактического материала выяснилось, как уже указывалось ранее, что в разновозрастных, но близких по составу отложениях иногда встречается больше одинаковых видов, чем в одновозрастных, но имеющих разный литологический состав.
| Рис. 12. Схема сопоставления отложений с применением процентно-статистического метода |
 |
В настоящее время этот метод для решения стратиграфических задач используют только в совокупности с другими палеонтологическими методами.
Несмотря на все преимущества палеонтологических методов, они не могут считаться универсальными. Немалый процент в составе земной коры составляют метаморфические,1 интрузивные и эффузивные породы, не содержащие ископаемые остатки организмов. Даже сред» осадочных пород нередко встречаются толщи, лишенные остатков организмов, или, как их называют, палеонтологически «немые». Для определения относительного возраста таких отложений палеонтологические методы неприменимы.
Дата добавления: 2019-12-09; просмотров: 646;
Поиск по сайту
Узнать еще
- I. Гидрометаллургические методы
- I. Погрешности механической обработки. Точность обработки. Методы их расчёта
- II. Методы исследования истории медицины.
- II. Пирометаллургические методы.
- II.II. Репродуктивные методы.
- II.III. Частично - поисковые или эвристические методы.
- II.V. Проблемные методы обучения.
- III. Методы изучения коллектива.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
