Модулирующие нейроны
В нервной системе выделена особая группа клеток – модулирующих нейронов, которые сами не вызывают реакции, но регулируют активность других нейронов. Они образуют контакты с другими нейронами типа «синапс на синапсе». Модулирующие нейроны причастны к регуляции болевой чувствительности. Схема взаимодействия модулирующего нейрона с сенсорным, проводящим сигналы о болевом раздражении, описана Л. Иверсон (1984) следующим образом. Синаптическая передача болевых сигналов в головной мозг, опосредуемая веществом Р, находится под контролем нейропептидов, подавляющих болевые сигналы. В задних рогах спинного мозга вставочные модулирующие нейроны, выделяющие нейропептид–энкефалин, образуют синапсы на аксонных окончаниях болевых нейронов. Энкефалин тормозит выход вещества Р, что уменьшает возбуждение постсинаптического нейрона, посылающего в головной мозг сигналы о боли.
Функции модулирующих нейронов наиболее хорошо изучены на моллюсках. Показана причастность модулирующих серотонинергических нейронов виноградной улитки к регуляции состояния организма. Улитку, которая ползала по шару, плавающему в воде, в течение 8 дней подвергали сильному электрическому раздражению вне сочетания с тестирующим стимулом – слабым тактильным раздражением. Это увеличивало величину реакции улитки на тактильный стимул – втягивание глазных щупалец. Эффект сохранялся в течение многих дней. Однако он проявлялся только в той обстановке, в которой применялось болевое раздражение (на шаре в воде) и отсутствовал, когда улитка тестировалась в других условиях – на стеклянной крышке аквариума. Применение нейротоксина, избирательно нарушающего работу серотонинергических нейронов, устраняло его проявление. Это одна из форм долговременной сенситизации, проявляющая себя как обстановочный условный рефлекс и требующая участия модулирующих серотонинергических нейронов. Предполагается, что данный обстановочный условный рефлекс реализуется на входах модулирующих нейронов, которые являются нейронами «состояния» [Балабан и др., 1992].
Модулирующие нейроны участвуют в процессе научения (см. гл. 15), изменяя проводимость синапса на пресинаптическом уровне. Эффект пресинаптической пластичности, создаваемый модулирующими нейронами, может быть начальным звеном в цепи событий, приводящих к пластической модификации поведения, связанной с выработкой условных рефлексов. Безусловное подкрепление как очень сильный раздражитель нейрона также возбуждает модулирующие нейроны и тем самым инициирует процесс пресинаптических изменений. За последним следует этап клеточных и молекулярных преобразований постсинаптического нейрона, характеризующий ассоциативное обучение.
Однако пресинаптическое облегчение, создаваемое модулирующими нейронами, может возникать и вне ситуации ассоциативного обучения только за счёт активации модулирующих нейронов сильными сенсорными раздражителями. Э. Кэндел и Р. Хокинс [1992] рассматривают его как результат «премодулирующего совпадения», которое на пресинаптическом уровне повышает проводимость, а само это повышение зависит от активности. Другое название этого явления – сенситизация, или неассоциативное обучение. Оно отличается от ассоциативного обучения, основанного на «пре-постсинаптическом совпадении» (рис. 8.1)
Эффект пресинаптического облегчения с участием модулирующих нейронов хорошо изучен при формировании оборонительного рефлекса у виноградной улитки. Внутриклеточная стимуляция её модулирующих нейронов, которые представлены пулом серотонинергических клеток педального ганглия, вызывает в командном нейроне оборонительной реакции появление деполяризационной волны и увеличение амплитуды суммарного ВПСП. Эти изменения превращают сенсорное подпороговое раздражение командного нейрона в пороговое, что выражается в появлении у него спайковой активности.
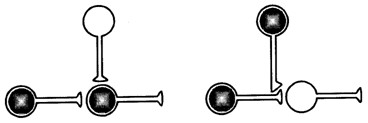
Рис. 8.1 . Два механизма изменения силы связи между нейронами. Один требует совпадения активности пресинаптического и постсинаптического нейронов во времени. Другой основан на совпадении активности пресинаптической клетки и модулирующей. Нейроны, участвующие в научении, заштрихованы [Кэндел, Хокинс, 1992]
Описаны два механизма длительного пресинаптического облегчения (фасилитации) под влиянием серотонина за счёт фосфорилирования двух типов ионных каналов: кальциевых и калиевых (рис. 8.2).
Участие кальциевых каналов в механизме повышения проводимости через синапс выглядит следующим образом. Во время потенциала действия ионы кальция и натрия входят в клетку, а ионы калия выходят из неё. Когда нейрон находится в активном состоянии, в нём увеличивается уровень содержания ионов кальция. Кальций, вошедший в клетку, действует на неё после того, как связывается с белком – кальмодулином. Этот комплекс так же, как и серотонин, активирует аденилатциклазу. За счёт двойного воздействия на неё резко увеличивается образование CAMP из аденозиттрифосфата (АТФ), который служит в клетке универсальным источником энергии для биохимических процессов; CAMP действует как вторичный посредник и активирует протеинкиназу. Протеинкиназа фосфорилирует рецепторные белки кальциевых каналов (присоединяет к ним фосфор). Это переводит кальциевые каналы в активное состояние – в клетку входит больше кальция, что увеличивает выделение из неё медиатора. Одновременно протеинкиназа фосфорилирует белки калиевых каналов. Это ведёт к уменьшению проводимости для ионов калия, т.е. к уменьшению калиевого тока, что ограничивает калиевую гиперполяризацию, следующую за ВПСП и потенциалом действия. Таким образом, потенциал действия продлевается, а это, в свою очередь, увеличивает продолжительность активного состояния кальциевых каналов.
Особую функцию в модуляции возбудимости нервной системы выполняют пейсмекерные нейроны (см. гл. 1). Будучи активированными, они своими длительными разрядами могут повышать проводимость сигнала через синапс. Специальный тип пейсмекерных нейронов, генерирующих разряды на частоте гамма-колебаний, которые, по-видимому, отражают колебания кальциевого тока на дендритах клеток таламуса, синхронизирует активность больших массивов нейронов, объединённых общими резонансными свойствами.
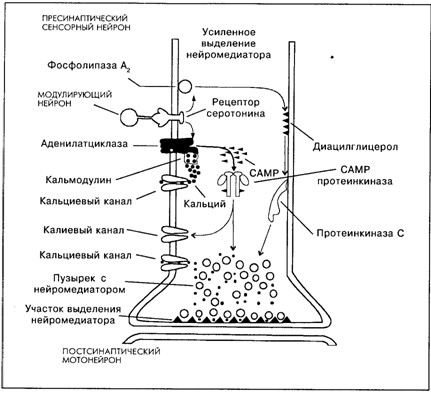
Рис. 8.2. Схема молекулярных реакций в пресинаптическом нейроне под влиянием возбуждения модулирующего серотонинергического нейрона – научение, зависимое от активности [Кэндел, Хокинс, 1992]
Дата добавления: 2016-06-05; просмотров: 1979;
Поиск по сайту
Узнать еще
- III и IV нейроны слухового пути. Третьи и четвертые нейроны слухового проводящего пути. Ядра слухового анализатора. Признаки поражения слухового пути.
- Вставочные нейроны.
- ГЛАВА 3. Мыслящее мясо: нейроны и синапсы
- Знаете ли вы? Делай, как я: зеркальные нейроны
- Клеточная нейроглия Нейроны
- Нейроны и глиальные клетки центральной нервной системы
- Нейроны и связи между ними

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
