F2: 9 сер. : 3 черн. : 4 бел.
Взаимодействие неаллельных генов.
План занятия.
1. Взаимодействие неаллельных генов.
2. Комплементарность.
3. Эпистаз.
4. Полимерия.
5. Плейотропное действие генов
6. Генотип — система генов.
1. Взаимодействие неаллельных генов.Очень редко развитие того или иного признака определяется одним геном. Часто один ген обусловливает развитие нескольких признаков. Такое явление называется плейотропным действием гена. Распространено и обратное — один признак контролируется несколькими генами.
Неаллельные гены— это гены, расположенные в различных участках (локусах)хромосом и кодирующие неодинаковые белки. Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием совокупности нескольких генов.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур.
Различные породы кур характеризуются разной формой гребня:
-Виандотты - низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”;
-Брамы и некоторые бойцовые куры - узкий и высокий гребень с тремя продольными возвышениями — “гороховидный”.
-Леггорны - простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному.
При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F2соответствует формуле 9 : 3 : 3 : 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
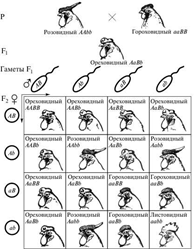 В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
1. Гибриды F1не похожи ни на одного из родителей и обладают новым признаком;
2. В F2появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Наследование формы гребня у кур можно отнести к комплементарному взаимодействию неаллельных генов.
2. Комплементарность.К комплементарным, или дополняющим друг друга, генам относятся такие неаллельные гены, которые при совместном проявлении обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.
Механизм комплементарного взаимодействияподробно изучен на примере наследования окраски глаз у дрозофилы.
Другой пример комплеменетарного действия генов наблюдается при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F2наблюдается расщепление в соотношении: 9 дисковидных : 6 сферических : 1 удлиненная (9:6:1)
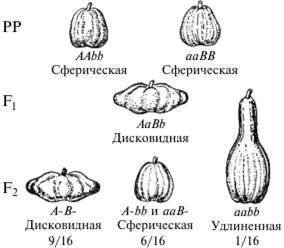 Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
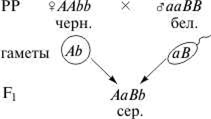
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ-), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb. Такое действие генов приводит к тому, что в F2расщепление по фенотипу соответствует формуле 9 : 3 : 4.
| AB | Ab | aB | ab |
| AB | AABB сер. | AABb сер. | AaBB сер. | AaBb сер. |
| Ab | AABb сер. | AAbb черн. | AaBb сер. | Aabb черн. |
| aB | AaBB сер. | AaBb сер. | aaBB бел. | aaBb бел. |
| ab | AaBb сер. | Aabb черн. | aaBb бел. | aabb бел. |
F2: 9 сер. : 3 черн. : 4 бел.
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F2наблюдается соотношение 3 : 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F1было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16 : 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.
Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.
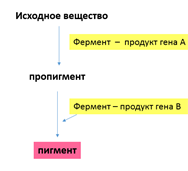 В приведенном примере формула расщепления в F2— 9 : 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление.
В приведенном примере формула расщепления в F2— 9 : 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление.
Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F1все гибриды имеют желтые зерновки, а в F2наблюдается расщепление 9/16 желт. : 7/16 фиол.
Таким образом, с позиции фенотипического проявления (расщепления по фенотипу в F2) можно выделить 4 варианта комплементарного взаимодействия генов.
1) 9 : 3 : 3 : 1- каждый из доминантных аллелей комплементарных генов в отдельности воспроизводит свой специфический признак (фенотип), а при совместном их присутствии в генотипе проявляется новый признак.
2) 9 : 6 : 1 — каждый доминантный аллель из комплементарных генов в отдельности воспроизводит один и тот же признак, а вместе они обусловливают проявление нового признака.
3) 9 : 3 : 4 - один из генов имеет собственное фенотипическое проявление, а действие другого может проявиться только в присутствии первого,что приводит к формированию нового признака.
4) 9 : 7 - каждый из комплементарных генов в отдельности не может обусловить развитие признака, но при сочетании в генотипе двух доминантных аллелей этих генов признак проявляется. При этом в F2 расщепление по фенотипу 9:7.
Эпистаз
Эпистаз— взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
Дата добавления: 2021-11-16; просмотров: 495;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
