Токсичность аммиака
1. повышение концентрации аммиака в мозге до 0,6 ммоль вызывает судороги.
2. аммиак в митохондриях сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глугамата:
3. повышение концентрации аммиака в крови сдвигает рН в щелочную сторону (вызывает алкалоз).
4. высокие концентрации аммиака стимулируют синтез глутамина из глутамата в нервной ткани (при участии глутаминсинтетазы):
5. накопление свободного иона NH4+ в цитозоле влияет на мембранный потенциал и работу внутриклеточных ферментов – он конкурирует с ионными насосами для Na+ и K+.
Обезвреживание аммиака
1. Системы безопасного транспорта аммиака по крови в органы обезвреживания:
· - глутаминовая система (перенос аммиака в печень и почки).
· - аспарагиновая система (перенос аммиака в печень)
· - глюкозо-аланиловый цикл (перенос аммиака из мышц в печень)
2. Защитный синтез мочевины в печени
3. Образование ионов аммония в почках
Биосинтез мочевины
- свободный аммиак и СО2 при участии 2АТФ образует макроэргическое соединение карбамоилфосфат (фермент - карбамоилфосфатсинтетаза). Карбамоилфосфат включается в орнитиновый цикл.
- Карбамоилфосфат отдает орнитину свою карбамоильную группу (фермент – орнитин-карбамоил-трансфераза) и образуется цитрулин и Н3РО4;
- Цитрулин взаимодействует с аспарагиновой кислотой с образованием аргининсукцината, при этом АТФ переходит в АМФ. Катализирует реакцию аргининсукцинатсинтаза;
- Аргининсукцинат расщепляется на фумарат и аргинин под действием аргининсукцинатлиазы;
- Аргинин под действием аргиназы (кофактор – ионы Ca или Mn) расщепляется путем гидролиза на мочевину и орнитин. Образующийся орнитин взаимодействует с новой молекулой карбамоилфосфата, и цикл замыкается.
Суммарное уравнение синтеза мочевины: СО2 + NH3 + Аспартат + 3 АТФ + 2 Н2О → Мочевина + Фумарат + 2 (АДФ + Н3Р04) + АМФ + H4P2O7.
Суточное выведение мочевины из организма составляет 20-35 гр. В норме концентрация мочевины в крови составляет величину 3,3-8,3 млмоль/л
Нарушение процессов обезвреживания аммиака приводит к его накоплению в крови (гипераммониемия).
Гипераммонеимия может быть: Первичной. В этом случае ее развитие обуславливается врожденной недостаточностью одного из ферментов мочевинообразования (карбомоилфосфатсинтетазы, орнитинкарбомоилтрансферазы).
Вторичной. Встречается при тяжелых поражениях печени.
6.4 Метаболизм нуклеотидов. Биосинтез и катаболизм пуриновых нуклеотидов. Источники атомов пуринового кольца. Нарушения обмена пуриновых нуклеотидов при подагре. Биосинтез и катаболизм пиримидиновых нуклеотидов. Источники атомов пиримидинового кольца. Гипероротатацидурия.
При синтезе нуклеотидов пуринового ряда, в отличие от синтеза пиримидиновых нуклеотидов, формирование гетероциклического ядра идет непосредственно на рибозо-5-фосфате. Вначале синтезируется ФРПФ, который при взаимодействии с глутамином превращается в 5-фосфорибозиламин:
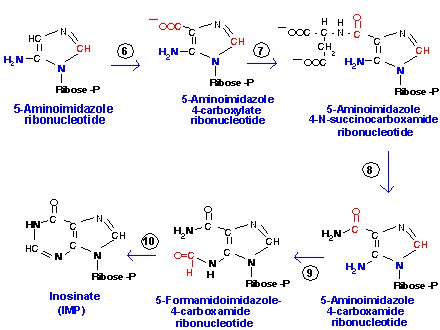
Затем следует большая последовательность реакций, в ходе которых формируется пуриновое ядро. Первым нуклеотидом, образующимся в ходе синтеза являетсяинозиновая кислота (ИМФ):
Источниками атомов углерода и азота при синтезе пуринового ядра являются следующие соедиения:
Глутамин, аспартат, глицин, углекислый газ образуются в организме, однако в условиях недостатка фолиевой кислоты могут возникнуть проблемы с обеспеченностью синтеза пуриновых нуклеотидов одноуглеродными группировками, переносчиками которых служит в клетках ТГФ.
Из ИМФ синтезируются другие нуклеотиды пуринового ряда. При синтезе АМФ идет аминирование ИМФ, источником аминогруппы служит аспартат.
При синтезе гуаниловой кислоты вначале остаток гипоксантина в ИМФ окисляется до ксантина с образованием КМФ, а затем идет аминирование и превращение КМФ в ГМФ. Донором аминогруппы выступает глутамин, энергетика реакции обеспечивается расщеплением АТФ. Образовавшиеся АМФ и ГМФ в ходе реакций трансфосфорилирования с АТФ преобразуются в АДФ и ГДФ, а затем последние подвергаются фосфорилированию за счет энергии, выделяющейся при биологическом окислении, превращаясь в АТФ и ГТФ.
Синтез пуриновых нуклеотидов с использованием в качестве пластического материала атомных группировок из молекул других соединений получил название синтеза “de novo”. В клетках млекопитающих работают также механизмы реутилизации азотистых оснований, образовавшихся в ходе внутриклеточного расщепления пуриновых нуклеотидов. Этот механизм синтеза пуриновых нуклеотидов получил название “синтез сбережения”.
Расщепление пуриновых нуклеотидовидет во всех клетках. Конечным продуктом катаболизма образующихся при расщеплении нуклеотидов пуриновых азотистых оснований является мочевая кислота. С наибольшей интенсивностью образование мочевой кислоты идет в печени, тонком кишечнике и почках.
Нуклеотиды в клетках подвергаются дефосфорилированию с образованием аденозина или гуанозина. Аденозин при участии фермента аденозиндезаминазы превращается в инозин и далее путем фосфоролиза в гипоксантин. Гипоксантин при участии ксантиноксидазы вначале окисляется в ксантин, а затем при участии того же фермента ксантин переходит в мочевую кислоту. При расщеплении ГМФ вначале в несколько этапов происходит образование свободного гуанина, который при участии фермента гуаназы переходит непосредственно в ксантин, а затем окисляется в мочевую кислоту.
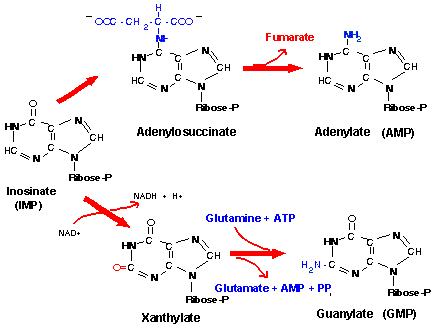
 Образовавшаяся мочевая кислота поступает в кровь и выводится через почки с мочой. Нормальное содержание мочевой кислоты в крови составляет 0,12–0,46 ммоль/л.
Образовавшаяся мочевая кислота поступает в кровь и выводится через почки с мочой. Нормальное содержание мочевой кислоты в крови составляет 0,12–0,46 ммоль/л.
Биосинтез нуклеотидов пиримидинового ряда начинается в цитозоле, где при участии цитозольной карбамоилфосфатсинтетазы образуется карбамоилфосфат, причем источником азота для его синтеза является глутамин:
СО2 + Глн + 2АТФ –® NH2-CO-O-PO3H2 + 2АДФ + Ф + Глу
Далее карбамоилфосфат взаимодействуя с аспартатом в реакции, катализируемой аспартаттранскарбамоилтрансферазой, превращается в карбамоиласпартат, а затем при участии дигигидрооротазы— в дигидрооротовую кислоту:
Дигидрооротовая кислота при участии митохондриального фермента дигидрооротатдегидрогеназы переходит в оротовую кислоту:
В следующей реакции принимает участие фосфорибозилпирофосфат. Он образуется из рибозо-5-фосфата с участием АТФ в ходе реакции, катализируемой ферментом фосфорибозилпирофосфатсинтетазой:
Реакция синтеза фосфорибозилпирофосфата (ФРПФ) не является специфичной для синтеза пиримидиновых нуклеотидов, в ходе этой реакции синтезируется ФРПФ, необходимый для синтеза различных мононуклеотидов.
Оротовая кислота при участии фермента оротат-фосфорибозилтрансферазы переносится на остаток рибозо-5-фосфата с образованием оротидиловой кислоты, которая подвергается декарбоксилированию, с образованием уридин-5-монофосфорной кислоты(уридиловая кислота или УМФ). Последняя реакция катализируется оротидилатдекарбоксилазой.
Все остальные нуклеотиды пиримидинового ряда синтезируются из уридиловой кислоты.
В ходе синтеза пиримидиновых нуклеотидов используются глутамин, СО2, АТФ, аспартат и фосфорибозилпирофосфат (ФРПФ). Все эти соединения синтезируются в клетках. Лишь при образовании из дУМФ дезокситимидиловой кислоты используется N5,N10-тетрагидрофолат; это значит, что при недостатке фолиевой кислоты (В9) в организме будет нарушен синтез дезокситимидиловой кислоты, необходимой для последующего синтеза ДНК в клетках.
свободные азотистые основания пиримидинового ряда повторно не используются и подвергаются расщеплению до конечных продуктов.
Расщепление пиримидиновых нуклеотидов начинается с отщепления рибозофосфатного остатка, а образовавшееся свободное азотистое основание расщепляется без образования специфических конечных продуктов.
Дата добавления: 2022-04-12; просмотров: 557;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
