Экстрапирамидные пути.
1) Красноядерноспинальный тракт. Первые нейроны располагаются в красном ядре среднего мозга. Их аксоны при выходе из покрышки среднего мозга совершают перекрест, после которого транзитом проходят через ретикулярную формацию моста, продолговатого мозга, спускаются в боковые канатики спинного мозга, доходят до своего сегмента и заканчиваются в двигательных ядрах передних рогов.
Здесь находятся тела вторых нейронов. Их аксоны в составе передних корешков выходят из спинного мозга и далее в составе спинномозговых нервов достигают поперечнополосатой мускулатуры.
2) Оливоспинальный тракт. Первые нейроны располагаются в нижней оливе продолговатого мозга. Их аксоны, не перекрещиваясь, идут в составе передних канатиков спинного мозга до двигательных нейронов передних рогов. Здесь располагаются тела вторых нейронов, чьи аксоны достигают скелетных мышц.
3) Вестибулоспинальный и ретикулоспинальный тракты. Первые нейроны располагаются в вестибулярных ядрах и ядрах ретикулярной формации, соответственно. Их аксоны, не перекрещиваясь, идут в составе передних канатиков спинного мозга до двигательных нейронов передних рогов. Здесь располагаются тела вторых нейронов, чьи аксоны достигают скелетных мышц.
4) Тектоспинальный тракт. Первые нейроны располагаются в четверохолмии среднего мозга. Их аксоны совершают перекрест, транзитом идут в передние канатики спинного мозга, и заканчиваются на двигательных нейронах передних рогов. Здесь располагаются тела вторых нейронов, чьи аксоны достигают скелетных мышц.
Функции нисходящих путей.Пирамидные и экстрапирамидные пути совместно обеспечивают регуляцию целенаправленных двигательных реакций. При этом функции пирамидной системы модулируются деятельностью экстрапирамидной системы.
Схема взаимодействия этих функциональных систем ЦНС выглядит следующим образом. При выполнении целенаправленного движения общее направление движения обеспечивается регуляторными механизмами экстрапирамидной системы. Она определяет пластический тонус мышц и обеспечивает сокращение мышечных групп, участвующих в движении. Завершение целенаправленного движения (например, захват предмета пальцами) осуществляется под контролем пирамидной системы и ее нисходящих путей. Нарушение взаимодействия пирамидной и экстрапирамидной систем при патологии ЦНС приводит к поражению двигательных реакций организма.
Экстрапирамидные пути осуществляют сложные безусловнорефлекторные реакции организма на раздражение. Например, тектоспинальный тракт обеспечивает ориентировочный рефлекс на неожиданные зрительные и слуховые раздражители; ретикулоспинальный тракт тормозит двигательные реакции спинного мозга.
Боковые желудочки мозга
Боковые желудочки являются полостью конечного мозга. Они залегают симметрично по обеим сторонам от средней линии ниже уровня мозолистого тела.
В каждом желудочке выделяют центральную часть, передний, задний и нижние рога.
Центральная часть соответствует теменной доле полушария и представляет собой горизонтально расположенное щелевидное пространство.
Передний рог бокового желудочка располагается в лобной доле, задний – в затылочной, нижний – в височной доле.
В центральную часть и нижний рог желудочка вдается сосудистое сплетение.
Сообщения желудочков головного мозга
1. Боковые желудочки через межжелудочковые отверстия сообщаются с III желудочком.
2. III желудочек, кроме вышеописанных боковых, сообщается через водопровод мозга с IV желудочком.
3. IV желудочек, кроме третьего, сообщается благодаря боковым и медиальному отверстиям с подпаутинным пространством головного мозга и с центральным каналом спинного мозга. Заращение этих отверстий ведет к накоплению жидкости в подпаутинном пространстве – водянке головного мозга.
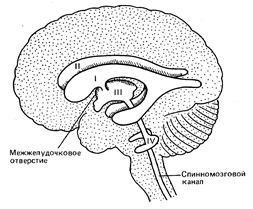
Желудочки головного мозга. Боковые желудочки (первый и второй) расположены в самой середине каждого полушария; третий желудочек занимает центральную часть большого мозга. Он сообщается с боковыми желудочками (через межжелудочковые, или монроевы, отверстия) и с четвертым желудочком, находящимся в стволе.
Оболочки головного мозга
Головной мозг, как и спинной, окружен тремя соединительнотканными оболочками: наружной твердой, средней паутинной и внутренней сосудистой.
Твердая оболочка. Ее наружная поверхность прилежит непосредственно к костям черепа, для которых является внутренней надкостницей. С костями свода черепа твердая оболочка связана слабо и легко отделяется, а с костями основания черепа – прочно.
По определенным линиям твердая оболочка расщепляется на два листка, образуя синусы (пазухи), выстланные эндотелием. В синусы впадают вены, по которым происходит отток крови из мозга. Основной отток крови из синусов идет через внутренние яремные вены. Синусы отличаются от обычных вен тем, что из-за наличия жестких стенок не спадаются и не изменяют свой просвет при изменениях внутричерепного давления.
Со своей внутренней стороны твердая оболочка образует отростки, проходящие в виде пластинок в щели между отдельными частями головного мозга и отделяющие эти части друг от друга.
Паутинная оболочка головного мозга имеет вид тонкой прозрачной, но плотной пластинки, бедной сосудами и нервами. От твердой оболочки она отделяется субдуральным пространством, а от мягкой – подпаутинным пространством. Подпаутинное пространство заполнено спинномозговой жидкостью. Паутинная оболочка соединяется с мягкой оболочкой с помощью соединитльнотканных перекладин.
Подпаутинное пространство не представляет общей полости одинаковой глубины, а состоит из множества сообщающихся между собой щелей. Там где подпаутинная оболочка перекидывается через глубокие борозды, образуются так называемые подпаутинные цистерны.
Все цистерны сообщаются друг с другом, а в области большого затылочного отверстия подпаутинное пространство головного мозга сообщается с подпаутинным пространством спинного мозга.
От наружной поверхности паутинной оболочки отходят специфические выросты, вдающиеся в венозные синусы твердой оболочки головного мозга. Это так называемые паутинные грануляции, через которые происходит отток спинномозговой жидкости из подпаутинного пространства в венозные синусы.
Мягкая (сосудистая) оболочка головного мозга плотно прилежит к веществу мозга. Она богата нервными и кровеносными сосудами и имеет большое значение в питании мозга. В определенных местах мягкая оболочка проникает в желудочки мозга и формирует складки, содержащие большое количество кровеносных сосудов. Это сосудистые сплетения, продуцирующие из крови спинномозговую жидкость.
Эмбриогенез спинного мозга
Нервная система плода начинает развиваться на ранних этапах эмбриональной жизни, продолжая развитие и в первые годы после рождения. Из эктодермы в заднем отделе зародыша образуется нервная пластинка, из которой впоследствии формируется нервный желобок, а затем - нервная трубка.
Из нее развивается спинной мозг, который соответственно сегментарному строению туловища делится на связанные между собою сегменты. Спинной мозг после 4 месяцев внутриутробного развития начинает отставать в росте от позвоночника. К рождению спинной мозг заканчивается на уровне третьего поясничного позвонка. У взрослого человека нижняя граница спинного мозга соответствует уровню 1-2 поясничных позвонков.
Спинной мозг у новорожденного имеет морфологически более зрелое строение, чем головной мозг. Его рост продолжается до 20 лет.
Эмбриогенез головного мозга
Головной мозг развивается из головного конца нервной трубки. Его преобразования тесно связаны с развитием органов чувств.
В эмбриогенезе головного мозга выделяют несколько стадий. На третьей неделе развития в головном отделе нервной трубки образуются три первичных мозговых пузыря (передний, средний и задний), из которых развиваются главные отделы головного мозга: конечный, средний и ромбовидный. В последующем передний и задний мозговые пузыри разделяются на два отдела, в результате чего образуется пять мозговых пузырей: конечный, промежуточный, средний, задний и продолговатый. Из конечного пузыря развиваются полушария головного мозга и подкорковые ядра; из промежуточного - промежуточный мозг (зрительные бугры, подбугорье, гипоталамус); из среднего - средний мозг (четверохолмие, ножки мозга); из заднего - мост и мозжечок, а из продолговатого - продолговатый мозг. К 3-му месяцу внутриутробного развития определяются основные части центральной нервной системы: большие полушария, ствол, мозговые желудочки, спинной мозг. К 5-му месяцу дифференцируются основные борозды коры больших полушарий, однако, кора остается еще недостаточно развитой.
Борозды и извилины головного мозга возникают благодаря неравномерному росту самого мозга. При этом выделяют первичные, вторичные и третичные борозды. Первичные борозды отличаются абсолютным постоянством. К ним относят, например, центральную, латеральную (боковую) и теменно-затылочную извилины. Латеральная борозда появляется к 4 месяцу внутриутробного развития, а теменно-затылочная и центральная – на 6-ом месяце.
Вторичные борозды появляются позднее (на 7-8 месяце внутриутробного развития) и характеризуются меньшим постоянством. Третичные борозды появляются не только в пренатальном, но и в постнатальном периоде развития (в течение 1-го месяца жизни ребенка). Это многочисленные мелкие борозды, непостоянные по времени появления, форме, числу и не имеющие названий.
Количество полушарных извилин, их форма, топографическое положение претерпевают определенные изменения по мере роста ребенка. Наибольшие изменения происходят в течение первых 5-6 лет, и лишь к 15-16 годам становятся похожими на мозговые структуры взрослого человека.
Головной мозг новорожденного имеет относительно большую величину, масса его в среднем составляет 1/8 массы тела. К моменту рождения вес мозга составляет 340 г у мальчиков и 330 г у девочек. К 9-месячному возрасту, первоначальная масса мозга удваивается и к концу первого года жизни составляет 1/11-1/12 массы тела. Наряду с ростом головного мозга меняются и ïðîïîðöèè черепа. Увеличение массы мозга происходит особенно интенсивно до 7-ми летнего возраста. Мозг достигает максимальной массы к 20-30 годам. У взрослого мужчины масса мозга – 1375 г, а у женщины – 1245 г. Причем масса мозга от периода рождения до взрослого возраста увеличивается в среднем в 4 раза, тогда как масса тела – в 20 раз.
Мозговая ткань новорожденного мало дифференцирована; корковые клетки и двигательные проводящие пути недоразвиты; вещество полушарий головного мозга слабо дифференцировано на белое и серое вещество. После рождения интенсивно протекают процессы миелинизации нервных волокон в различных отделах головного мозга. К 9-ти месяцам миелинизация в большинстве волокон коры больших полушарий достигает хорошего развития, за исключением коротких ассоциативных волокон в лобной доле. К этому возрасту становятся более отчетливыми первые три слоя коры.
Миелинизация волокон, расположение слоев коры и дифференциация нервных клеток в основном завершается к 3 годам.
В младшем школьном возрасте, а также в период полового созревания продолжающееся развитие головного мозга характеризуется увеличением количества ассоциативных волокон и образованием новых нервных связей.
Функциональные особенности развития коры. У новорожденного большие полушария головного мозга не обладают регулирующим влиянием на нижележащие отделы ЦНС. Кора не регулирует движения, поэтому у новорожденного они имеют обобщенный характер и не имеют целенаправленности, за исключением движений, обеспечивающих прием пищи. Считается, что на ранних этапах периода новорожденности функции ребенка регулируются в основном промежуточным мозгом. Рефлекторные дуги безусловных рефлексов проходят через зрительные бугры и полосатое тело.
Развитие периферической нервной системы
Периферическая нервная система новорожденного недостаточно миелинизирована. Миелинизация проходит неравномерно. Так черепномозговые нервы миелинизируются в течение 3-4 мес., процесс заканчивается к 1 году 3 мес. Миелинизация спинномозговых нервов продолжается до 2-3 лет.
Вегетативная нервная система функционирует у ребенка с момента рождения. После рождения отмечается лишь образование отдельных узлов и мощных сплетений симпатической нервной системы.
Рассмотренные данные свидетельствуют о том, что уже на самых ранних стадиях эмбриогенеза развития нервной системы осуществляется по принципу системогенеза с развитием в первую очередь тех отделов, которые обеспечивают жизненно необходимые врожденные реакции, создающие первичную адаптацию ребенка после рождения (пищевые, дыхательные, выделительные защитные).
Изучение развивающегося мозга, особенно в первый год жизни, обнаруживает нечто сходное: появление новых форм реагирования сопровождается угасанием, редукцией первичных автоматизмов. Но при этом оба эти процесса должны быть сбалансированы. Преждевременное угасание первичных автоматизмов лишает функции прочного фундамента, так как при развитии мозга принцип преемственности обязателен. В то же время слишком поздняя редукция устоявшихся форм реагирования мешает образованию новых, более сложных реакций.
Сбалансированность процессов редукции и обновления наиболее выступает в двигательном развитии детей первого года жизни. Так, при рождении у ребенка имеются первичные позотонические автоматизмы, влияющие на мышечный тонус в зависимости от положения головы в пространстве. К концу второго - к началу третьего месяца жизни эти автоматизмы должны угасать, уступая место новым формам регуляции мышечного тонуса - способности ребенка удерживать голову. Если этого не происходит, данные автоматизмы следует рассматривать как аномальные, ибо они препятствуют удерживанию головы. Формируется патологическая связь: невозможность удерживать голову нарушает развитие зрительного восприятия и вестибулярного аппарата; из-за неразвитости вестибулярного аппарата не вырабатывается способность к распределению тонуса мышц, обеспечивающему акт сидения. И как итог - искажается вся схема двигательного развития.
Таким образом, наряду с гетерохронностью развития отдельных функциональных систем и их звеньев необходима и определенная синхронность их взаимодействия. Для каждого возрастного периода отдельные системы должны иметь определенную зрелость, иначе не произойдет нормального слияния систем в единый ансамбль.
Вегетативная нервная система
Вегетативную нервную систему подразделяют на симпатическую и парасимпатическую. Работа этих двух систем-антагонистов поддерживает в организме стабильность внутренней среды перед лицом вечно изменяющегося мира.
Главное различие между симпатической и парасимпатической системами заключается в том, что первая мобилизует организм для действия (катаболизм), а другая - восстанавливает запасы энергии в организме (анаболизм).
Основная функция симпатической системы - это мобилизация всего организма при чрезвычайных, экстремальных обстоятельствах. Такая мобилизация связана с рядом сложных реакций, начиная с расщепления гликогена в печени (образующаяся при этом глюкоза служит добавочным источником энергии) и кончая изменениями в циркуляции крови. Каждую из этих реакций, осуществляемых симпатической нервной системой, легко понять как механизм приспособления к «аварийным» ситуациям, выработанным в ходе эволюции. Обеспечение доступа к запасам энергии дает организму максимум физических возможностей в непредвиденных ситуациях. Уменьшение кровотока около поверхности тела снижает вероятность обильного кровотечения при повреждении кожи, тогда как усиленная подача крови к глубже лежащим мышцам позволяет развить большее физическое усилие. Кеннон назвал весь этот комплекс изменений «реакцией борьбы и бегства». Его теоретические соображения о роли этой реакции явились существенным стимулом для развития психофизиологии и современных представлений об «общей активации» организма.
Следующее по важности различие - то, что симпатическая система имеет тенденцию действовать быстро и как единое целое, тогда как парасимпатическая активация более кратковременна и носит локальный характер. Действие симпатической системы обычно проявляется диффузно (охватывает весь организм) и поддерживается относительно долго. С другой стороны, действие парасимпатической системы, способствующее сохранению и поддержанию основных ресурсов организма, локально и относительно кратковременно.
Эффекты симпатической и парасимпатической систем на органы и системы организма противоположны друг другу. В то время как симпатическая нервная система ускоряет сокращения сердца, парасимпатическая их замедляет, она усиливает также приток крови к желудочно-кишечному тракту и стимулирует превращение глюкозы в гликоген печени. Большинство, но не все внутренние органы получают иннервацию от обеих систем.
Поскольку обе они работают согласованно, трудно бывает определить, связано ли данное изменение функции с активностью той или другой из них. Например, замедление ритма сердца может указывать и на усиленную активность парасимпатической системы, и на ослабление действия ее антагониста.
Последнее различие в функционировании обеих систем связано с особенностями их структурной организации. В соматической нервной системе каждый нейрон, тело которого находится в ЦНС, имеет длинный отросток – аксон, проводящий нервные импульсы к органу-мишени. В произвольной мускулатуре такой аксон образует синапс в области двигательной пластинки мышечного волокна. Соматическая нервная система, таким образом, имеет «однонейронный путь». В вегетативной же системе путь к органу-исполнителю двухнейронный. Место соединения между этими двумя нейронами находится в вегетативном ганглии.
Симпатические волокна выходят из средней части спинного мозга - из грудного и поясничного отделов, поэтому симпатическую систему иногда называют тораколюмбальной. Ее аксоны сходятся к группе симпатических ганглиев, расположенных с обеих сторон спинного мозга. В этих ганглиях с плотно расположенными нейронами существуют большие возможности для электрических «переключений», и импульс, пришедший из любого участка симпатической системы, может вызвать активацию всей этой системы. Парасимпатические волокна образуют синапсы недалеко от иннервируемого органа, они выходят из спинного мозга выше или ниже места выхода симпатических волокон - из черепного и кресцового отделов. В связи с этим парасимпатическую нервную систему еще называют краниосакральной. Ее ганглии расположены далеко друг от друга, и поэтому нервные импульсы оказываются более специфическими. Кроме этого симпатическая нервная система иннервирует надпочечники, которые при стимуляции выделяют адреналин и норадреналин. Норадреналин гормонального происхождения попадает в симпатические синапсы и усиливает их действие.
Для распада этого гормона требуется некоторое время, поэтому для прекращения активности симпатической системы также требуется некоторое время. Для парасимпатической системы существует другой медиатор - ацетилхолин. В синапсах этой системы ацетилхолин быстро инактивируется холинэстеразой, в связи с этим парасимпатические эффекты четко ограничены не только в пространстве, но и во времени. Важным исключением из этого правила являются симпатические волокна, иннервирующие потовые железы - они активируются ацетилхолином. Таким образом, различия между катаболическим действием симпатической системы и анаболическим действием парасимпатической системы обусловлены особенностями их анатомического строения.
Нервные механизмы регуляции вегетативных функций имеют иерархическую структуру. Первым уровнем этой иерархии являются внутриорганные периферические рефлексы, замыкающиеся в интрамуральных ганглиях вегетативной нервной системы, средним уровнем являются нервные центры ствола мозга, высшим уровнем - кора больших полушарий. Она обеспечивает координацию вегетативных и соматических функций в сложных поведенческих реакциях организма, возникновение которых обусловлено индивидуальным опытом.
Установлено, что в продолговатом мозге расположены центры, тормозящие деятельность сердца, возбуждающие слезоотделение и секрецию слюнных и желудочных желез, поджелудочной железы, вызывающие выделение желчи из желчного пузыря и желчного протока, возбуждающие сокращение желудка и тонкого кишечника. Здесь же, в ретикулярной формации находится сосудодвигательный центр, координирующий и интегрирующий деятельность нейронов симпатического отдела нервной системы, расположенных в тораколюмбальных сегментах спинного мозга и посылающих на периферию сосудосуживающие импульсы.
Характерной особенностью сосудодвигательного центра продолговатого мозга и нейронов блуждающего нерва, тормозящих сердечную деятельность, является то, что они находятся постоянно в состоянии автоматической активности (тонуса), в результате чего артерии и артериолы всегда несколько сужены, а сердечная деятельность замедлена.
При участии нейронов ядер блуждающих нервов осуществляется различные рефлексы на сердце, в том числе рефлекс Гольца, глазосердечный (рефлекс Ашнера), дыхательно-сосудистый и др.
Многие рефлекторные реакции сердца осуществляются сопряженно с изменениями сосудистого тонуса. Это обусловлено связями, существующими между нейронами, регулирующими деятельность сердца и сосудистый тонус.
Импульсы к спинномозговым нейронам симпатической системы, иннервирующим сосуды, передаются от сосудовигательного центра по ретикулоспинальным путям. Сосудорасширяющие рефлексы сосудодвигательного центра имеют, как правило, регионарный характер, сосудосуживающие - охватывают обширные области тела.
В отличие от дыхательного центра, центры регуляции сердечной деятельности и сосудистого тонуса, хотя и находятся под влиянием коры полушарий мозга, обычно не могут быть произвольно возбуждены или заторможены (для этого требуется специальная тренировка).
Рефлекторные центры продолговатого мозга, регулирующие деятельность пищеварительных органов, осуществляют свое влияние через парасимпатические нервные волокна, приходящие к слюнным железам в составе языкоглоточного и лицевого нервов, а к желудку, поджелудочной железе, тонкому кишечнику, желчному пузырю и желчным протокам - в составе блуждающего нерва.
В среднем мозге, в передних буграх четверохолмия, находятся нервные центры зрачкового рефлекса и аккомодации глаза.
Деятельность вегетативных центров, расположенных в спинном, продолговатом и среднем мозге, в свою очередь регулируется высшими вегетативными центрами гипоталамуса.
Особенность ответных реакций, возникающих при раздражении разных участков гипоталамуса, заключается в том, что в них участвуют многие органы тела. Эти реакции являются комплексными и интегрированными. Ядра гипоталамуса принимают участие во многих общих, в том числе поведенческих, реакциях. Так, гипоталамус участвует в половых и агрессивно-оборонительных реакциях. Точечное раздражение его вентромедиального ядра вызывает у кошки резко выраженный агрессивный эффект - так называемую реакцию мнимой ярости.
Деятельность гипоталамуса в свою очередь контролируется высшими отделами ЦНС - подкорковыми ядрами, мозжечком и корой больших полушарий, с которыми гипоталамус связан как прямыми нервными путями, так и путями, проходящими в составе ретикулярной формации мозгового ствола.
Оказывая активирующее и тормозящее влияние на различные отделы ЦНС, ретикулярная формация повышает активность вегетативных нервных центров. Она оказывает на них тонизирующее влияние. Поэтому симпатический отдел может рассматриваться в функциональном единстве с ретикулярной формацией.
Экспериментально показано, что введение адреналина повышает тонус ретикулярной формации, в результате чего усиливается ее активирующее влияние на большие полушария.
На вегетативную нервную систему существенное влияние оказывает мозжечок. При его удалении возникает угнетение моторной, в частности периодической, деятельности пищварительного тракта и секреторной функции желез желудка и кишечника.
Как известно, подкорковые ядра, в частности полосатое тело, участвуют в осуществлении сложных, безусловно-рефлекторных реакций организма, которые включают и вегетативные компоненты. Вегетативные реакции могут формироваться при возбуждении подкорковых ядер вследствие того, что последние имеют прямые связи с ретикулярной формацией мозгового ствола и гипоталамусом.
Важную роль в регуляции деятельности внутренних органов имеют нервные образования, которые входят в состав лимбической системы, или висцерального мозга: гиппокамп, поясная извилина, миндалевидные ядра.
Лимбическая система участвует в формировании эмоций и таких поведенческих реакций, в осуществлении которых имеет место ярко выраженный вегетативный компонент. Влияние висцерального мозга на функции органов, иннервируемых вегетативной нервной системой, осуществляется через гипоталамус.
Высшим уровнем регуляции вегетативной нервной системы является кора полушарий мозга. Впервые В.Я. Данилевским в 1874 г. было установлено, что раздражение некоторых участков коры больших полушарий у собак вызывает изменения дыхания, сердечной деятельности, сосудистого тонуса. Впоследствии В.М. Бехтерев, Н.А. Миславский, другие физиологи и нейрохирурги наблюдали изменения многих вегетативных функций при раздражении разных участков коры больших полушарий.
Школа И.П. Павлова рассматривает нейроны коры больших полушарий, участвующих в регуляции функций внутренних органов, как корковое представительство интероцептивного анализатора. В регуляции вегетативных функций большое значение имеют лобные доли коры больших полушарий. Раздражение этих долей вызывает изменение дыхания, пищеварения, кровообращения и половой деятельности. Поэтому считается, что в передних отделах коры больших полушарий находятся высшие центры вегетативной нервной системы.
В коре больших полушарий существуют зоны, связанные нисходящими путями с ретикулярной формацией ствола мозга. Эти зоны расположены в сенсомоторной коре, лобных глазодвигательных полях, поясной извилине, верхней височной извилине и околозатылочной области. По нисходящим путям, идущим от этих зон коры, импульсы поступают к ретикулярной формации, а от нее - к гипоталамусу и гипофизу. Имеются также прямые пути, идущие от лобной доли и от поясной извилины к гипоталамусу. С помощью этих путей кора оказывает регулирующее влияние на нижележащие уровни вегетативной нервной системы.
Вегетативная нервная система регулирует работу сердца, желез и непроизвольной (гладкой) мускулатуры без активного участия нашего сознания. В течение многих лет считалось, что функции вегетативной системы недоступны для нормального самоконтроля. Недавние эксперименты с созданием обратной связи и изучение практики восточных мистиков с их древней религиозной традицией тренировки тела позволяют предполагать, что и так называемую «непроизвольную» мускулатуру можно поставить под контроль воли.
Влияние коры головного мозга на многие внутренние органы доказано в опытах с воздействием на человека гипнотического внушения. Например, показано, что внушением можно вызвать учащение или замедление деятельности сердца, расширение или сужение сосудов, усиление отделения мочи почками, выделение пота, изменение интенсивности обмена веществ.
Известны случаи, когда влияние коры полушарий мозга проявлялось настолько резко, что человек мог произвольно вызывать увеличение частоты сердечных сокращений, поднятие волос и появление «гусиной» кожи, обычно наблюдаемой в результате охлаждения тела, а также изменять ширину зрачков, зависящую от тонуса гладких мышц радужки глаза.
Однако, если бы протекание внутренних функций требовало нашего повседневного контроля, у нас почти не оставалось бы времени на что-либо иное, кроме непрерывных забот о поддержании жизнедеятельности собственного организма. Специализация и автоматический режим работы вегетативной нервной системы создали возможность для дальнейшей эволюции мозговых механизмов, обеспечивающих высшие психические функции.
Электрическая активность мозга (электроэнцефалография)
Мозг состоит из более чем 10 млрд клеток и каждая из них представляет собой миниатюрную станцию, способную в возбужденном состоянии создавать электрический потенциал. Впервые эта электрическая активность была зарегистрирована в 1875 г. английским хирургом Ричардом Ксйтоном. Он впервые показал, что у животного можно зарегистрировать электрическую активность мозга. В своих экспериментах этот исследователь регистрировал фоновую активность с поверхности сенсорной коры мозга кролика.
Приблизительно через 50 лет (1924) сходные наблюдения были сделаны на человеке австрийским психиатром Хансом Бергером. Запись мозговых волн осуществлялась с помощью специального прибора, способного регистрировать и усиливать потенциалы, создаваемые нервными клетками. Это делалось с помощью электродов, прикрепленных к коже черепа испытуемого. Регистрируемые слабые потенциалы усиливались и отображались графически в виде волн, которые Бергер назвал "мозговые волны". В ходе своих исследований он установил, что часть этих потенциалов принадлежит мозгу, а не обусловлена активностью мышц головы; что электрические характеристики этих сигнапов зависят от состояния испытуемых; он выделил основные виды электрической активности мозга: апьфа-волны и бета-волны; показал что электрическая активность мозга может рассматриваться в качестве индикатора общей активности мозга. Метод регистрации электрической активности мозга получил название электроэнцефалографии (ЭЭГ).
Последующие исследования показали, что показатели ЭЭГ качественно отличаются от открытых ранее более простых показателей активности вегетативной нервной системы. Периодические волны электрической активности, возникающие при сокращении сердца, - это сама простота по сравнению с ужасающей сложностью ЭЭГ. Сложность формы волн ЭЭГ как будто бросает вызов исследователям, пытающимся найти в них хоть какой-нибудь смысл.
Современники Бергера отнеслись к его сообщениям скептически и "мозговые воины" стали общепризнанным фактом только после того, как Эндриан и Мэттьюз осуществили наглядную демонстрацию записи ЭЭГ на заседании английского физиологического общества в 1935 г.
Последующие годы были волнующими для исследователей, которые с энтузиазмом проникали в тайны глубин мозга. Многочисленные работы позволили установить:
т.тектро энцефалографические корреляты эпилепсии:
записи с разных участков черепа региетрирх ют колебания разной формы:
- при опухолях мозга в окружающей их ткани обычно появляются аномальные медленные волны и ЭЭГ можно использовать для уточнения локализации таких опухолей:
- ЭЭГ позволяет судии, о функциональном состоянии коры, например, о глубине наркоза, о наличии и определенных зонах патологических процессов.
Было проведено бесчисленное количество исследований, направленных на поиски ЭЭГ-коррелятов интеллекта, особенностей личности, поведения. Результаты в большинстве случаев оказались разочаровывающими. В мозгу человека более 10 млрд нервных клеток, сплетенных в плотную есть взаимными связями. Даже в самых гонких записях ЭЭГ неизбежно выявляется лишь слитная трескотня сотен тысяч клеток, приглушенная и искаженная черепом. По мнению одного из исследователей: "Мы подобны слепым, пытающимся понять работу фабрики, прислушиваясь снаружи к се шуму" (Margcrison ct al., 1967).
Изучение изменений ЭЭГ начинается с анализа частот и амплитуд мозговых волн.
- альфа-ритм: при низкой активности мозга большие группы нервных клеток разряжаются одновременно. Эта синхронность отображается на ЭЭГ в виде последовательности медленных волн с частотой от 8-13 гц, имеющих большую амплитуду. Альфа-ритм наиболее четко регистрируется главным образом в затылочных отведениях, когда человек находится в расслабленном состоянии, с закрытыми глазами.
- бета-ритм: во время активной работы мозга каждая участвующая в ней нервная клетка разряжается в соответствии со своей специфической функцией в своем собственном ритме. В результате активность становится абсолютно асинхронной и регистрируется в виде быстрых волн высокой частоты (выше 13 Гц) и малой амплитуды. Амплитуда бета-волн уменьшается по мере увеличения активности мозга. Ранние наблюдения о связи этих волн с состоянием относительной активности были в общем подтверждены.
- тета-ритм: колебания от 4 до 8 гц. Они появляются на первой стадии сна, а также у некоторых опытных мастеров медитации или во время пребывания испытуемых в изолированной камере в условиях сенсорной депривации.
- дельта-ритм: колебания с частотой менее 4 гц. Регистрируется но время глубокого сна, а также при некоторых патологических состояниях (опухоли мозга) или у больных незадолго до смерти.
Следует подчеркнуть, что разбиение на группы по частоте более или менее произвольно - оно не соответствует каким-то физиологическим категориям. И связь со степенью "психической активации" представляется довольно слабой, так как есть много исключений. Напимер, у взрослых людей при эмоциональных переживаниях обнаруживаются колебания гста-частоты. Кроме того, эта классификация не подходит для всех возрастов: альфа-ритм появляется только в раннем подростковом возрасте.
При первых попытках более систематического анализа изменений ЭЭГ обычно измеряли и частоту, амплитуду. Одна из наиболее часто используемых зависимых переменных - это "время альфа-ритма", т.е. процент времени, занимаемый альфа-ритмом.
Еще один традиционный показатель ЭЭГ - это "блокада альфа-ритма", т.е. внезапное очень резкое уменьшение амплитуды альфа-волн, которое обычно происходит при предъявлении раздражителей. Например, если на глаза испытуемого падает свет, то в затылочных отделах мозга у него обычно происходит блокада альфа-ритма.
Появление компьютерной техники способствовало появлению новых методов вычисления различных показателей ЭЭГ. В частности, многие исследователи стали проводить частотно-амплитудный анализ регистрируемых волн электрической активности мозга. По мере развития компьютерных способов математической обработки ЭЭГ появлялись вес новые возможности для изучения механизмов работы мозга.
Так, электроэнцефалографические исследования Л.П. Павловой (1988) обнаружили для разных видов деятельности человека универсальное правило "смещения фокуса максимальной активации (ФМА). Например, при выполнении сознательных целенаправленных действий ФМА устойчиво регистрируется в речевых зонах левого полушария. Затем возникает генерализация возбуждения и распространение ФМА на ряд других корковых зон. Другими словами, наблюдается смена доминантно-субдомннантных отношений между полушариями и лобными и теменными ассоциативными областями коры. Завершающаяся перестройка активности коры сводится к устойчивому смещению ФМА в правое полушарие, а затем в задние отделы коры. В лобных долях на этой стадии ФМА наблюдается в течение коротких промежутков времени. Наиболее выраженное смещение ФМА обнаружено в процессе предметной (внешнеразвернутой) деятельности и в процессе мыслительной (свернутой) деятельности. Интеллектуальные психические процессы всегда в
Дата добавления: 2018-11-26; просмотров: 601;
Поиск по сайту
Узнать еще
- Верхнее строение пути.
- Вопрос 30. Теоретическое определение замедления и тормозного пути.
- Задача о maxmin пути.
- Лекция 15. И.А.Гончаров (1812-1891). Начало творческого пути.
- Определение потребной тормозной силы по заданной длине тормозного пути.
- Первые искусственные водные пути. Вышневолоцкая система.
- Порядок действий локомотивной бригады и работников, связанных с движением поездов, в случае обнаружения неисправности – «толчок» в пути.
- Проверки и осмотр пути.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
