II. Травматическая регенерация.
1. Регенерация, обусловленная дедифференцировкой клеток:
а) заживление ран.
Эпидермис и первичная кора дедифференцируются, их клетки начинают делиться и образуют вторичную меристему, которая превращается в пробку.
б) органогенез, связанный с образованием каллуса.
Клетки дедифференцируются и переходят к неорганизованному делению, образуя каллусную ткань из рыхло соединенных друг с другом паренхимных клеток. Иногда отдельные клетки дают начало адвентивным, то есть возникшим не из эмбриональных тканей, органам: корням, побегам, листьям.
в) соматический эмбриогенез.
На раневой поверхности образуется каллус. Из отдельных клеток каллуса, начинающих делиться, формируются соматические зародыши (эмбриоиды), из которых при определенных условиях развивается целый организм.
г) восстановление частей без образования каллуса.
Паренхимные клетки коры под влиянием ауксина, индуцирующего генетическую программу ксилемообразования, превращаются в клетки ксилемы при образовании обходного участка проводящего пучка вокруг места его прерывания.
2). Регенерация на уровне меристем:
а) восстановление апикальных меристем.
При продольном рассечении конуса нарастания из каждой половины могут регенерировать отдельные апексы.
б) органогенез из предсуществующих зачатков.
Восстановление надземных органов у высших растений происходит за счет отрастания пазушных почек при устранении доминирующего влияния апекса побега.
10.5. Кинетика ростовых процессов
Кривую, описывающую скорость роста, можно разделить на 4 участка: 1) лаг-период, когда рост почти не заметен и идут процессы, подготавливающие организм к видимому росту, 2) лог-фаза, когда скорость роста изменяется логарифмически, 3) фаза замедления роста, 4) стационарная фаза (рис. 10.1).
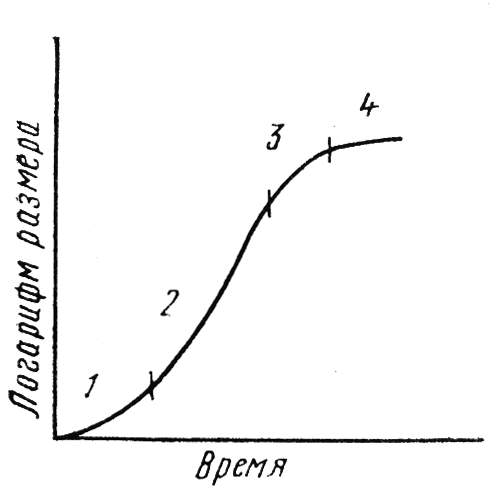
Рис. 10.1. Кривая роста.
1 – лаг-период, 2 – логарифмическая фаза, 3 – фаза замедленного роста, 4 – фаза стационарного состояния (по С. И. Лебедеву).
Для измерения скорости роста используются следующие показатели.
Удельная скорость роста r – прирост массы растения или отдельного его органа в единицу времени, который рассчитывается по формуле Блекмана:
W1
lg ¾ x 2,3026
W0
r = ¾¾¾¾¾¾¾¾
t
где W0 – начальный, а W1 – конечный вес сухого вещества, t – промежуток времени между определениями.
Относительный или процентный рост R– прирост, вычисленный в процентах от исходного веса растения или органа: (W1 – W0)
R = ¾¾¾¾ x 100
W0
Абсолютная скорость роста К– величина прироста за промежуток времени, отнесенная к единице времени:
W2 – W1
K = ¾¾¾
t2 – t1
10.6. Влияние факторов внешней среды на рост растений
На рост растений оказывают влияние продукты жизнедеятельности других растений (явление аллелопатии), микроорганизмов (антибиотики, регуляторы роста) и факторы внешней среды.
Свет.Растения воспринимают свет не только как источник энергии, но и в качестве сигнала, характеризующего условия среды. В клетках имеются рецепторные молекулы фитохрома, опосредующие действие света на морфогенез. Фитохром состоит из двух белковых субъединиц и хромофора – незамкнутого тетрапиррола, относящегося к группе фикобилинов. Фитохром синтезируется в форме Ф660, поглощающей красный свет. Под действием красного света он переходит в активную форму Ф730, поглощающей дальний красный свет. Под действием дальнего красного света и в темноте Ф730 превращается в Ф660. Фитохром изменяет проницаемость клеточных мембран, регулирует движение хлоропластов и влияет на синтез ферментов и стимуляторов роста гиббереллинов и цитокининов.
Температура.Различают три основные температурные точки: минимальная температура, при которой начинается рост, оптимальная – наиболее благоприятная для роста и максимальная, при которой рост прекращается. В зависимости от приспособленности к температурному режиму различают теплолюбивые (минимальная температура выше 10оС, оптимальная 30-40оС) и холодостойкие (минимальная температура 0-5оС, оптимальная 25-30оС).
Газовый состав.Необходим кислород, так как дыхание поставляет энергию для ростовых процессов, и углекислый газ, который в ходе фотосинтеза восстанавливается до органических веществ. Избыток углекислого газа на короткое время повышает растяжимость клеточных стенок и стимулирует рост клеток (эффект «кислого роста»).
Водный режим.Недостаточное снабжение растений водой задерживает рост побегов и кратковременно стимулирует с последующим торможением рост корней.
Минеральное питание.Для нормального роста необходимо достаточное снабжение всеми питательными элементами. Избыток азота стимулирует рост вегетативной массы, но замедляет процессы дифференцировки и формирование цветков.
10.7. Фитогормоны
Они образуются в процессе обмена веществ растений и оказывают в очень малых количествах регуляторное и координирующее влияние на физиологические процессы в разных органах растения. Различают стимуляторы и ингибиторы роста. Стимуляторы роста, применяемые в сверхоптимальных дозах, способны подавлять ростовые процессы.
10.7.1. Ауксины
Главным представителем ауксинов в растениях является индолил-3-уксусная кислота (ИУК). Она синтезируется из триптофана в верхушке побега. Разрушается ИУК ферментом ИУК-оксидазой. Ауксин стимулирует деление и растяжение клеток, необходим для образования проводящих пучков и корней. ИУК активирует протонную помпу в плазмалемме, что приводит к закислению и разрыхлению клеточной стенки и тем самым способствует росту клеток растяжением. Комплекс ИУК с рецептором транспортируется в ядро и активирует синтез РНК, что в свою очередь приводит к усилению синтеза белков.
10.7.2. Цитокинины
Цитокинины образуются путем конденсации аденозин-5-монофосфата и изопентенилпирофосфата в апикальной меристеме корня. Много цитокининов в развивающихся семенах и плодах. Цитокинины индуцируют в присутствии ауксина деление клеток, активируют дифференциацию пластид, повышают активность АТФ-синтетазы, способствуют выходу почек, семян и клубней из состояния покоя, предотвращают распад хлорофилла и деградацию клеточных органелл. Ткани, обогащенные цитокининами, обладают высокой аттрагирующей способностью. Комплекс цитокининов с белковым рецептором повышает активность РНКполимеразы и экспрессию генов. При этом увеличивается число полисом и активируется синтез белка.
10.7.3. Гиббереллины
В настоящее время известно более 70 гиббереллинов кислой и нейтральной природы. Наиболее известным и распространенным гиббереллином является гибберелловая кислота. Гиббереллины синтезируются из ацетилкоэнзима А в листьях и корнях. Гиббереллины способствуют удлинению стебля, выходу семян из состояния покоя, формированию гранулярного эндоплазматического ретикулума, образованию цветоноса и цветению, активируют деление клеток в апикальных и интеркалярных меристемах, повышают активность ферментов синтеза фосфолипидов. Комплекс гиббереллина с белковым цитоплазматическим рецептором стимулирует синтез нуклеиновых кислот и белка.
10.7.4. Абсцизовая кислота
Она синтезируется в листьях и корневом чехлике двумя путями: из мевалоновой кислоты или путем распада каротиноидов. Абсцизовая кислота (АБК) тормозит рост растений и является антагонистом стимуляторов роста. Однако АБК активирует удлинение гипокотиля огурца, образование корней у черенков фасоли. АБК ускоряет распад нуклеиновых кислот, белков, хлорофилла, ингибирует мембранную протонную помпу. АБК накапливается в клетках при неблагоприятных условиях внешней среды, стареющих листьях, покоящихся семенах, в отделительном слое черешков листьев и плодоножек.
10.7.5. Этилен
Газ этилен синтезируется из метионина или путем восстановления ацетилена. Много его накапливается в стареющих листьях и созревающих плодах. Он ингибирует рост стеблей и листьев. Удлинение стебля тормозится из-за изменения направления роста клеток с продольного на поперечное, что приводит к утолщению стебля. Обработка этиленом индуцирует корнеобразование, ускоряет созревание плодов, прорастание пыльцы, семян, клубней и луковиц.
10.7.6. Брассиностероиды
Брассиностероиды содержатся в разных органах растений, но особенно много их в пыльце. Они стимулируют рост в длину и толщину проростков, усиливая как деление, так и растяжение клеток.
10.7.7. Синтетические регуляторы роста
Ретарданты ингибируют рост стебля благодаря торможению растяжения клеток и подавлению синтеза гиббереллинов. Стебли становятся более короткими и утолщаются, в результате повышается устойчивость растения к полеганию.
Морфактиныпрепятствуют прорастанию семян, образованию и росту побегов, ослабляют апикальное доминирование у побегов и усиливают его у корней.
Гербицидыслужат для уничтожения растительности. Есть гербициды общего действия, когда погибают все растения, и селективные для избирательного уничтожения определенных классов растений. Они могут подавлять фотосинтетическое или окислительное фосфорилирование.
Дефолиантыускоряют листопад у растений, что активирует созревание семян и плодов и облегчает механизированную уборку урожая.
Десикантывызывают ускоренное высушивание листьев и стеблей, что позволяет вести сбор семенников бобовых культур и уборку картофеля комбайнами.
Сениканты– смесь физиологически активных веществ, вызывающих ускорение созревания и старения сельскохозяйственных растений.
ДВИЖЕНИЯ РАСТЕНИЙ
Движение – это перемещение организма или его частей в пространстве. Движение необходимо для питания, защиты и размножения растений.
11.1. Верхушечный рост
Перемещение корневых волосков и гиф грибов происходит за счет верхушечного роста. В кончиках клеток содержится много везикул, производных аппарата Гольджи и эндоплазматического ретикулума, которые доставляют материалы для формирования клеточной стенки, а также синтетазы и гидролазы. Гидролазы разрыхляют стенку, которая растягивается под давлением протопласта. Микрофибриллы целлюлозы раздвигаются и в стенку встраиваются новые компоненты.
11.2. Ростовые движения
В процессе эволюции у растений возник специфический способ движения за счет необратимого роста клеток растяжением.
11.2.1. Тропизмы
Это движения растений, обусловленные односторонне действующими факторами среды. Они являются следствием более быстрого роста клеток растяжением на одной стороне побега, корня, листа. В зависимости от природы фактора различают гео-, фото-, тигмо-, термо-, электро- и хемотропизмы. При положительных тропизмах движение направлено в сторону раздражителя, при отрицательных – от него. Рассмотрим несколько видов тропизма.
Геотропизм– изгибание под действием гравитационного поля Земли вследствие разной скорости роста клеток противоположных сторон органа. Восприятие силы тяжести связано с воздействием на плазмалемму и эндоплазматический ретикулум статолитов (амилопласты, хлоропласты, отложения солей). Давление статолитов на мембраны приводит к изменению мембранного транспорта, поляризации клеток. При этом нижняя часть стеблей и корней, приведенных в горизонтальное положение, приобретает суммарный электроположительный заряд. Активируется транспорт ауксина как отрицательно заряженного аниона к нижней стороне органа. У стебля это приводит к усилению роста нижней части горизонтально расположенного стебля и его изгиба вверх. У клеток корней выше чувствительность к ауксину, поэтому повышенная концентрация ИУК в нижней части корня ингибирует ее рост и верхняя половина растет быстрее, что приводит к изгибу корня вниз.
Фототропизм– изгибы растений под влиянием одностороннего освещения. Затененная сторона этиолированных проростков растет более быстро, так как туда перемещается ауксин. В зеленых проростках ауксин не транспортируется на затененную часть побега, но у них в клетках освещенной стороны накапливается ингибитор роста ксантоксин.
Хемотропизм– ростовая реакция на химические соединения. Он наблюдается у корней, пыльцевых трубок, железистых волосков насекомоядного растения росянки. Большую роль при этом играет не только природа вещества, но и его концентрация: при низкой концентрации хемотропизм положителен, а при сверхоптимальной – отрицателен.
11.2.2. Ростовые настии
Это движения растений, обусловленные диффузно действующими факторами среды и происходят в результате неравномерного роста клеток растяжением. При росте верхней стороны орган изгибается книзу (эпинастия), нижней стороны – кверху (гипонастия).
Фотонастиявызвана сменой света и темноты. У многих растений цветки закрываются при наступлении темноты и открываются утром. Эти движения можно вызвать, искусственно затеняя и освещая растения. Движения цветков происходят из-за разной скорости роста верхней и нижней сторон лепестков при изменении освещенности.
Тигмонастияхарактерна для лазящих растений, когда усик в ответ на прикосновение закручивается. Он реагирует только при создании эффекта трения. В темноте усик не закручивается, но освещение растения даже через 90 мин после прикосновения вызывает закручивание усика. Движение происходит при участии этилена и ауксина, так как обработка ими вызывает закручивание усика без механического раздражения. После прикосновения нижняя сторона усика теряет тургор и становится вогнутой. Затем начинает расти верхняя сторона усика, что приводит к его закручиванию.
11.2.3. Круговые нутации
Это круговые или колебательные движения органов растения, характерные, в основном, для растущих побегов и корней. Они происходят за счет идущих по кругу ускорений роста клеток в результате геотропической корректировки в гравитационном поле.
11.3. Тургорные обратимые движения
Сюда относят движения замыкающих клеток устьиц, которые мы уже изучали. Такие же движения характерны для растений, чьи листья имеют специализированные образования в сочленениях между стеблем и черешком, между черешком и пластинкой листа. В этих образованиях паренхимные клетки коры, изменяя свой тургор, приводят к складыванию и разгибанию листьев. Различают флексорные и экстензорные клетки. Повышение тургора во флексорных клетках приводит к складыванию листьев, в экстензорных – к их разгибанию. Изменения тургора в этих клетках связано с изменениями транспорта сахарозы, ионов калия и хлора в результате деятельности протонных помп. Эти движения могут происходить медленно при смене условий освещения (никтинастические или движения сна), при механическом и химическом воздействиях у росянки или с большой скоростью при сотрясении у мимозы, кислицы и других растений.
Дата добавления: 2016-07-11; просмотров: 2140;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
