Митохондриальная ДНК
К настоящему времени прочитано более 100 разных геномов митохондрий. Набор и количество их генов в митохондриальных ДНК (мтДНК), для которых полностью определена последовательность нуклеотидов, сильно различаются у разных видов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomonas americana (97 генов). У большинства высших животных геном митохондрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК). У растений и простейших, в отличие от животных и большинства грибов, в митохондриальном геноме закодированы и некоторые белки, входящие в состав рибосом этих органелл. Ключевые ферменты матричного полинуклеотидного синтеза, такие как ДНК-полимераза и РНК-полимераза, зашифрованы в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерархии эукариотической клетки.
Геномы митохондрий разных видов отличаются не только по набору генов, порядку их расположения и экспрессии, но по размеру и форме ДНК. Подавляющее большинство описанных сегодня митохондриальных геномов представляет собой кольцевые суперспирализованные двуцепочечные молекулы ДНК.
Как правило, в каждой митохондрии содержится несколько копий ее генома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них по 10 одинаковых геномов.
Две нуклеотидные цепи молекулы мтДНК имеют различный суммарный нуклеотидный состав и обозначаются как L-цепь (light, легкая) и Н-цепь (heavy, тяжелая). Различия обусловлены неодинаковым содержанием "тяжелых" пуриновых и "легких" пиримидиновых нуклеотидов. Большая часть молекулы митохондриальной ДНК содержит консервативные кодирующие последовательности. Только небольшой не кодирующий участок размером 1122 нуклеотидные пары, так называемый "контрольный регион", содержит гипервариабельные участки HV1 (342 пары нуклеотидов), HV2 (268 пар нуклеотидов) и HV3 (137 пар нуклеотидов) (рис. 11.1).
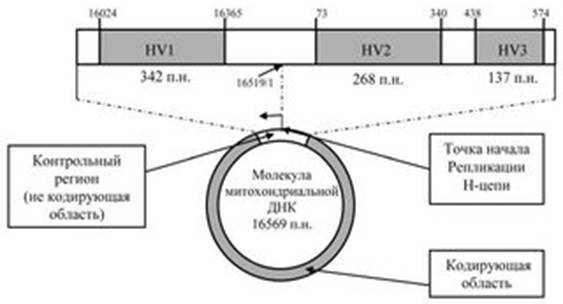
Рисунок 11.1. Участки митохондриальной ДНК человека
Большая часть генов расположена на Н-нити мтДНК: это 14 генов тРНК, оба гена рРНК и 11 генов, кодирующих белки. 8 тРНК генов и два белок-кодирующих гена (ND3 и АТ8)расположены на L-нити (рис. 11.2). Малый размер митохондриального генома животных по сравнению с грибами и растениями при сохранении почти всех митохондриально кодируемых белков указывает на его чрезвычайно экономную организацию. Так, гены расположены почти вплотную, межгенные спейсеры составляют всего до 9 нуклеотидов.
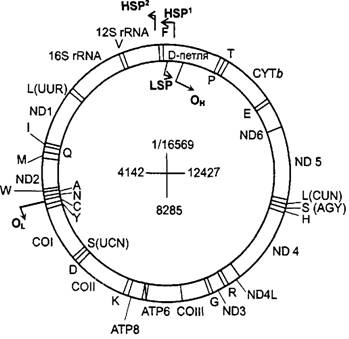
Рисунок 11.2. Карта митохондриальной ДНК человека. Указаны гены субъединиц NAD-дегидрогеназы, цитохрома b, субъединиц цитохрома с, субъединиц АТФ-синтазы, рРНК (12S и 16S) и тРНК (однобуквенные символы аминокислот). Стрелками указаны промоторы транскрипции цепей (LSP, HSP1, HSP2) и точки инициации репликации (0L, Он).
Имеющихся в митохондриальной ДНК 22 тРНК генов достаточно для синтеза всех необходимых митохондриальных белков. 13 генов мтДНК животных кодируют белки различных энергетических комплексов. Гены митохондриальных белков почти всегда фланкированы с двух сторон генами тРНК. Единственным протяженным участком мтДНК животных, не имеющим никаких генов, является область так называемой D-петли (Displacement loop). Эта структура, видимая в электронный микроскоп, состоит из двуцепочечного и одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок формируется частью L-цепи и комплементарным ей вновь синтезированным фрагментом ДНК длиной 450 - 650 нуклеотидов, имеющим на 5'-конце рибонуклеотидную затравку, которая соответствует точке начала синтеза Н-цепи (ori H). Синтез L-цепи начинается лишь тогда, когда дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область инициации репликации L-цепи доступна для ферментов синтеза ДНК лишь в одноцепочечном состоянии, а, следовательно, только в расплетенной двойной спирали при синтезе Н-цепи. Таким образом, дочерние цепи мтДНК синтезируются непрерывно и асинхронно (рис. 11.3).
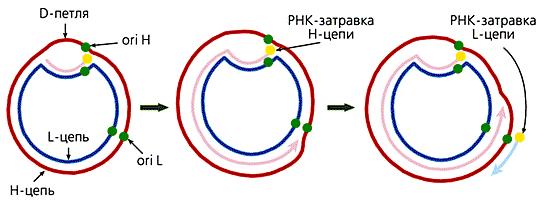
Рисунок 11.3 Схема репликации мтДНК млекопитающих
Длина D-петли несколько различается у разных видов и определяется короткими (15 п.н.) специфическими TAS-последовательностями (termination-associated sequences).
ДНК-репликация в митохондриях тесно связана с транскрипцией. В области D-петли расположены два промотора: один из них (HSP) инициирует транскрипцию основной кодирующей Н-цепи, другой (LSP) - комплементарной легкой L-цепи. В области D-петли находится также точка инициации репликации Н-цепи (Он), тогда как точка инициации репликации L-цепи находится обычно вдали от D-петли.
Вначале на LSP инициируется синтез коротких РНК-транскриптов, которые служат праймерами для начала репликации Н-цепи. Вновь синтезируемая часть Н-цепи имеет небольшую длину (до 1000 п.н.). Она комплементарно связывается с соответствующим участком L-цепи, оттесняя в этом месте родительскую Н-цепочку, так что образуется D-петля. Молекула мтДНК в области D-петли трехнитчатая.
Митохондриальная ДНК человека имеет все признаки, характерные для мтДНК млекопитающих: типичный размер - 16569 п.н, кольцевую структуру, 37 генов, реплицируется с образованием D-петли и экспрессируется с помощью ряда ядерных факторов.
Наряду с мономерными кольцами в митохондриальных геномах человека и других млекопитающих редко встречаются димерные и мультимерные молекулы, которые особенно характерны для малигнизированных клеток.
Кроме механизма асинхронной репликации Н- и L-нитей мтДНК в митохондриях млекопитающих обнаружена синхронная репликация двух цепей, начинающаяся в точке Он и продолжающаяся в одном направлении по всей длине молекулы. Данный способ репликации характерен для клеток, восстанавливающих нормальное число молекул мтДНК после резкого их уменьшения, тогда как асинхронная репликация типична для клеток, поддерживающих число копий мтДНК при стабильном состоянии
В отличие от большинства эукариотических генов, которые транскрибируются независимо друг от друга, каждая из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК, начинающейся в районе ori H. Помимо этих двух длинных молекул РНК, комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые начинаются в той же точке и заканчиваются на 3'-конце гена 16S рРНК (рис. 11.4). Таких коротких транскриптов в 10 раз больше, чем длинных. В результате процессинга из них образуются 12S рРНК и 16S рРНК, участвующие в формировании митохондриальных рибосом, а также фенилаланиновая и валиновая тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются транслируемые мРНК, к 3'-концам которых присоединяются полиадениловые последовательности. 5'-концы этих мРНК не кэпируются, что необычно для эукариот. Сплайсинга не происходит, поскольку ни один из митохондриальных генов млекопитающих не содержит интронов.
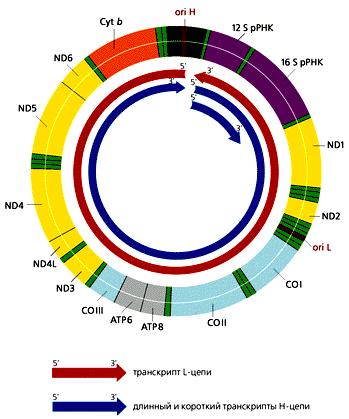
Рисунок 11.4. Транскрипция мтДНК человека
Доля молекул с D-петлей в общем пуле митохондриальных ДНК зависит от типа клеток и стадии клеточного цикла. Так, при увеличении потребности в кислороде частота молекул мтДНК с D-петлей увеличивается.
В митохондриях общее число молекул с D-петлей значительно превышает число полностью реплицирующихся молекул. Обусловлено это тем, что у D-петли есть дополнительные функции - это прикрепление мтДНК к внутренней мембране и инициация транскрипции, поскольку в этом районе локализованы промоторы транскрипции обеих цепей ДНК.
Высокая мутабильность митохондриальной ДНК связана, с одной стороны, с особенностями ее организации: геном не защищен гистоновыми белками, репарационные процессы не так многообразны, как в ядре. В то же время митохондрии поглощают более 90% кислорода, попадающего в клетку, и дыхательная цепь образует при функционировании большое количество ДНК-повреждающих свободных радикалов. Высокая концентрация активных форм кислорода в митохондриях увеличивает частоту мутаций мтДНК по сравнению с ядерной на порядок. Радикалы кислорода служат причиной специфических замен Ц-Т (дезаминирование цитозина) и Г-Т (окислительное повреждение гуанина), вследствие чего, возможно, мтДНК богаты АТ-парами. Кроме того, все мтДНК обладают интересным свойством - они не метилируются, в отличие от ядерных и прокариотических ДНК.
Аккумулируя точечные мутации почти в десять раз быстрее, чем ядерная, мтДНК любого представителя незамкнутой популяции отличается от мтДНК другого человека в среднем на 25 нуклеотидных замен, которые не всегда являются патологическими, а часто представляют результат полиморфизма. Этот полиморфизм создает объективные трудности в выяснении истинной роли мтДНК в патогенезе, его следует учитывать при любом исследовании митохондриального генома.
Генотип человека по локусам митохондриальной ДНК называется митотипом. Для обозначения аллельных вариантов, при описании митотипа, указывается отличие последовательности нуклеотидов аллеля конкретного митотипа от стандартной последовательности. В качестве стандартной последовательности нуклеотидов молекулы митохондриальной ДНК человека используется так называемая пересмотренная Кембриджская референсная последовательность (revised Cambridge Reference Sequence или rCRS) опубликованная в 1999 году.
Работы Брауна с соавторами (Brown et al., 1980; 1982), посвященные сравнению последовательностей мтДНК у приматов и человека, у представителей различных человеческих рас показали высокую информативность мтДНК, как инструмента исследований эволюционной, популяционной и исторической генетики. Этому способствуют уникальные особенности мтДНК, во-первых, высокая скорость изменчивости мтДНК приводит к высокому уровню полиморфизма мтДНК в популяциях. Во-вторых, отсутствие рекомбинаций и наследование мтДНК исключительно гаплотипами способствует простоте реконструкции предковых форм – гаплотипов-основателей – наблюдаемого генетического разнообразия; и, наконец, наследование от одного родителя определяет мтДНК как нейтральный маркер материнской линии.
В настоящее время наиболее распространены два основных метода и подхода к исследованию полиморфизма мтДНК в популяциях мира. Первый подход – это высокоразрешающее рестрикционное картирование тотальной молекулы мтДНК. Работы в этом направлении выявили высокий уровень разнообразия митохондриального генофонда и показали наличие этно-географической специфичности типов мтДНК. На основе диагностических рестрикционных сайтов в различных, преимущественно кодирующих, участках мтДНК типы мтДНК объединяются в континент-специфические группы, или гаплогруппы, описывающие от 60 до 100% общего разнообразия последовательностей мтДНК населения отдельных континентов.
Второй подход – это исследование полиморфизма главной некодирующей области (D-петли) мтДНК. Полиморфизм первого гипервариабельного сегмента D-петли может на данный момент считаться наиболее изученным в популяционном плане. Несмотря на большое разнообразие индивидуальных линий мтДНК показано, что эти гаплотипы дифференцируются на группы. Так для населения Европы и Ближнего Востока выявлено пять таких гаплогрупп, объединяющих близкородственные гаплотипы на основе наиболее устойчивых ассоциаций нуклеотидных замен.
На сегодня уже установлено, какие митохондриальные линии присущи населению Западной и Восточной Евразии, Африки, Австралии и Америки. По современным представлениям, все евразийские группы мтДНК входят в состав трех макрогрупп - M, N и R, которые произошли примерно 65 тыс. лет назад из африканской митохондриальной группы L3. Распределение линий мтДНК в генофонде населения Евразии характеризуется выраженной этнорасовой специфичностью. Например, генофонды народов западной и восточной частей Евразии различаются кардинально. Почти все линии мтДНК корейцев и бурят относятся к набору восточноевразийских групп - A, B, C, D, F, G, M7, M8a, M9, M10, M11, Y, Z, N9a, R9. Европейцам же свойственны группы H, HV*, pre-V, T, J, K, U2, U3, U4, U5, U8, N1a, I, W и X. В другой части Евразии, в северной лишь в краевых популяциях почти полностью преобладают митохондриальные линии одного этнорасового происхождения - монголоидного или европеоидного. Огромная же по протяженности территория (от Алтая до Восточно-Европейской равнины) представляет собой древнюю контактную зону. Именно в ней проходил расогенез за счет межрасового смешения (рис. 11.5).
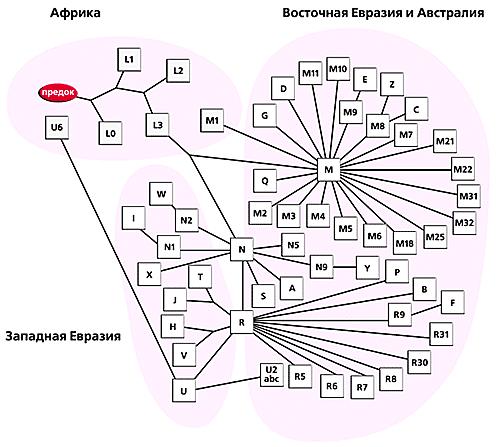
Рисунок 11.5. Филогенетическое дерево митохондриальной ДНК человека
Митохондриальный генофонд русского населения характеризуется высоким разнообразием, но встречаются и общие группы. Наиболее частыми из них были H, U, T и J - те же группы, которые широко распространены в генофондах других европейских народов. Монголоидная примесь у русских оказалась незначительной - около 1.5%, причем ее составляли группы мтДНК (C, D, M*, G2a, N9a) восточноевразийского происхождения (рис. 11.6).
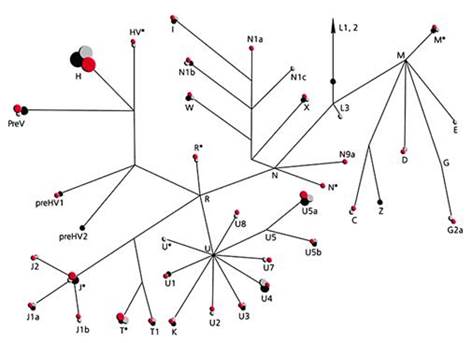
Рисунок 11.6. Структура митохондриального генофонда русских, поляков и боснийцев (кластеры мтДНК обозначены буквам (розовыми кружками (их размер пропорционален частоте гаплотипа мтДНК) даны кластеры русских, серыми - поляков, черными – боснийцев).
Дата добавления: 2022-07-20; просмотров: 247;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
