ХАРАКТЕРИСТИКИ ВЕЛИЧИНЫ И СТРУКТУРЫ ПОПУЛЯЦИИ
Популяция имеет определенную организацию и структуру, которые в той или иной степени поддаются описанию. Организация популяции подразумевает в первую очередь такие ее характеристики, как общая величина, характер распределения особей внутри популяции (включая плотность распределения) и механизмы, обеспечивающие устойчивое поддержание популяции как самостоятельной единицы. Под структурой популяции в широком смысле понимают любые подразделения популяции как единичного целого на связанные в определенном порядке части.
2.1. Величина популяции
Величина популяции может характеризоваться в двух аспектах: по числу особей, составляющих популяцию, и по пространству, ею занимаемому (ареалу популяции). Определить первую величину можно только зная вторую. В сравнительно редких случаях, имея дело с очень небольшими популяциями, их величину можно определить путем простого подсчета особей. Гораздо чаще приходится прибегать к так называемому приему экстраполяции: определив среднюю плотность популяции умножать ее на площадь всего ареала, рассчитывая таким образом приблизительное число особей всей популяции.
Очень часто исследователь, пытающийся определить на практике величину конкретной популяции, сталкивается с ситуацией, когда внутри вида, представленного множеством популяций, бывает трудно узнать границы конкретной популяции. Между тем определение таких границ зачастую представляет и значительный практический интерес в связи с пониманием популяции как единицы управления биологическим разнообразием. В связи с этим в рамках популяционной биологии во второй половине ХХ в. сформировалось особое направление исследований – биохорология, устанавливающая подходы и методы определения границ реальных природных популяций. Многообразие экологических связей и особенности филогенетической истории различных биологических видов определили весьма широкий спектр различий в отношении величины их популяций и степени выраженности их пространственных границ.
В природе не так уж редко встречаются популяции с относительно небольшими по площади и весьма четко очерченными ареалами. К числу таковых относятся, в частности, реликтовые популяции, оставшиеся от распространенной в прошлом флоры или фауны. Например, включенный в Красную книгу Республики Беларусь козелец голый (Scorzonera ruprechtiana) на территории нашей страны сохранился только в одном месте – на крутом склоне холма возле оз. Глубелька (национальный парк «Нарочанский»). Небольшая популяция этого растения, безусловно, давно отделена от других популяций данного вида, локализованных на севере Уральских гор. К числу реликтовых относятся также популяции нескольких видов так называемых ледниковых реликтовых ракообразных, изолированно обитающих в нескольких озерах севера
Беларуси: лимнокалянус (Limnocalanus macrurus), реликтовая мизида (Mysis relicta), бокоплав Палласа (Pallasea quadrispinosa), понтопорея (Pontoporeia affinis). Немало других примеров такого рода нетрудно найти в Красной книге любой страны.
Не представляет особых сложностей и определение границ островных популяций, под которыми подразумеваются не только популяции собственно островов, где в качестве фактора изоляции выступают море или океан. К этому же типу относятся и популяции, ареалы которых четко очерчены сушей. Например, популяции рыб гольцов (Salvelinus) в озерах Сибири могут быть настолько хорошо дифференцированными, что особи из разных озер нередко сильно отличаются друг от друга. Хорошо изолированными ареалами обладают очень многие виды высокогорных животных, характеризующиеся ограниченной подвижностью (жужелицы, моллюски и др.) и относящиеся к альпийской зоне. Некогда единый ареал таких популяций оказался раздроблен на ряд фрагментов, разделенных пониженными межгорными котловинами и реками и вполне подходящих под характеристику островных популяций.
В настоящее время многие большие ареалы популяций раздроблены на ряд островных ареалов из-за деятельности человека. В нашей стране это явление происходит с глухарем (Tetrao urogallus). Из-за неграмотной эксплуатации популяций данного вида охотничьих птиц, а также сокращения площади пригодных биотопов и ряда других причин, ареал глухаря в Беларуси на сегодня представлен серией изолированных друг от друга территорий, разделенных пространствами, на которых этой птицы нет.
На фоне общей низкой численности данного оседлого вида существует реальная опасность генетического вырождения многих его островных популяций. У других организмов популяции, напротив, занимают гигантские по площади ареалы, очертить достаточно точные границы которых непросто. Наиболее показательные примеры здесь демонстрируют животные, характеризующиеся высокой степенью подвижности. Например, у европейской городской ласточки (Delichon urbica) все особи, населяющие Европу и Западную Сибирь, представляют по сути единую популяцию.
Их концентрация на зимовке в экваториальной Африке способствует сохранению единого генофонда, и ласточка, появившаяся на свет и окольцованная в гнезде, например, в предместьях Лондона, на следующий год может гнездиться в Новосибирске.
Однако и у перелетных птиц, совершающих ежегодные миграции на большие расстояния, ареал порой оказывается дифференцирован на ряд самостоятельных популяций. В таких случаях именно разные районы зимовок способствуют поддержанию самостоятельности нескольких популяций, например у красноголовой чернети (Aythya ferina), у которой в пределах Евразии выделяются четыре популяции, зимующие в Западном Средиземноморье, бассейне Нила, Индии и Юго-восточной Азии. При этом, как видно из рис. 1, гнездовые ареалы этих популяций отчасти перекрываются, и в некоторых районах бок о бок могут гнездиться особи двух соседних популяций.
Подобные зоны перекрывания ареалов популяций доставляют объективные трудности при их картировании, создавая впечатление отсутствия дискретности популяции как таковой. Однако изучение объективной картины распределения особей в пространстве с учетом их экологической и фенотипической специфики обычно позволяет наметить достаточно выраженные границы популяций и определить зоны их перекрывания. Определенным подспорьем при решении подобных задач может стать использование таксономической категории подвида. Правда, как уже отмечалось выше, подвиды выделяются далеко не во всех группах организмов. «Традиционным» их выделение является у животных с хорошо исследованной географической изменчивостью – у позвоночных, отдельных отрядов насекомых и некоторых других групп. Подвиды представляют собой группы популяций, объединяемых по набору каких-либо морфологических признаков. Чем более ярко выражены такие признаки (например, вариации рисунка крыла у бабочек, окраски оперения у птиц и т. п.) и чем более явственно привязаны их особенности к конкретным географическим регионам, тем легче выделяются подвиды. Однако и здесь исследователи порой сталкиваются с проблемой перекрывания подвидовых ареалов, в которых особи разных подвидов гибридизируют, создавая фенотипическую картину интерградации («перехода») признаков. Определение границ ареалов подвидов, таким образом, может рассматриваться как промежуточный этап в определении и картировании границ ареалов конкретных популяций.
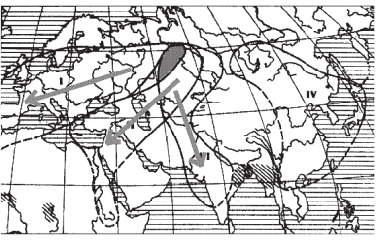
Рис. 1. Популяции красноголовой чернети (Aythya ferina) в Евразии (стрелками показаны направления осенних миграций разных популяций; цветом выделен участок перекрывания ареалов трех популяций в Западной Сибири)
Изучение экологии популяций традиционно велось главным образом на животных; именно в применении к животным были заложены и разработаны существующие подходы в определении границ популяций и подвидов. Однако в последнее время интенсивно стали исследоваться и популяции растений, по отношению к которым ботаники часто употребляют термин «ценопопуляция», который подчеркивает место и роль определенного вида растений в функционировании фитоценоза.
Растительная ценопопуляция включает все особи вида в пределах данного фитоценоза независимо от их фонетического состояния, экотопических и генетических особенностей. Такой подход к характеристике популяций растений определил достаточно четкий принцип выделения конкретных ценопопуляций; их размер и границы определяются границами фитоценоза, которые, хотя и нелинейны, но морфологически выражены более четко, чем границы популяций у животных. Сложности с выделениями границ популяций существуют и в этом случае, но они связаны не со специфическими популяционными проблемами, а с проблемами классификации растительности и методов выделения фитоценозов.
2.2. Пространственное распределение особей в популяции
Внутри ареала популяции особи редко распространяются равномерно. Различают три типа пространственного распределения: случайное (диффузное), равномерное (регулярное) и агрегированное (мозаичное) (рис. 2).
Случайное распределение в природе встречается нередко; в этом случае расстояния между ближайшими особями в популяции различны. Это связано как с неоднородностью среды обитания популяции, так и, в значительной степени, со случайными явлениями. Данный тип распределения часто отмечается у растений и многих животных – в частности у тех, у кого социальные связи между особями выражены относительно слабо. Равномерное распределение характеризуется равным удалением каждой особи от всех соседних особей. В природе такое распределение встречается достаточно редко: на участках с однородной средой обитания, в частности в моновидовых зарослях некоторых растений и плотных поселениях некоторых сидячих беспозвоночных.
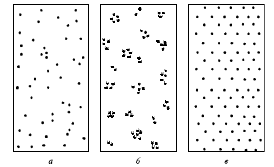
Рис. 2. Типы пространственного распределения особей в популяции: а - случайное; б - групповое (агрегированное); в - равномерное
Агрегированное распределение выражается в образовании группировок особей, между которыми остаются достаточно большие незаселенные участки. Биологически это связано либо с резкой неоднородностью среды, либо с выраженной социальной структурой внутри популяции, как, например, у животных, образующих колонии, либо у растений – потомков вегетативного размножения одной особи.
Пространственное распределение особей часто отражает те или иные особенности состояния популяции: так, при максимально возможной плотности на пригодных участках нередко наблюдается регулярное распределение особей, а если популяция далека от предельной насыщенности – распределение бывает случайным. Агрегированное распределение часто наблюдается, если пригодные для жизни вида участки в пределах ареала популяции «вкраплены» среди непригодных территорий.
Как величина популяции, так и характер распространения особей внутри нее зависят не только от распределения пригодных для жизни условий, но и от биологических особенностей организмов, в первую очередь их подвижности. В биологических исследованиях разных групп организмов нередко возникает необходимость в определении характеристик подвижности особей того или иного вида либо популяции. В частности, исследования перемещений птиц и некоторых других животных с применением мечения и кольцевания позволяют определить среднюю величину дисперсии – среднего расстояния от места появления на свет до места первого размножения особей.
В некоторых случаях удалось выявить даже межпопуляционные различия средней дисперсии в разных популяциях одного вида. Так, исследования обыкновенных поползней (Sitta europaea), населяющих широколиственные леса Центральной Европы, показали, что среднее расстояние между местом появления на свет и местом первого гнездования окольцованных птенцами особей составляет всего лишь около 800 м, а максимальное не превышает 27 км, тогда как у популяций данного вида, гнездящихся в Западной Сибири, эти величины в 2 раза выше.
Однако даже в приведенном примере видно довольно большое различие между величинами средней и максимальной дисперсии, что создает определенные неудобства. Во многих отношениях более показательной характеристикой подвижности особей в популяции является радиус репродуктивной активности (РРА). В настоящее время в популяционной экологии закрепилось понимание этой величины, основанное на эмпирических данных: радиусом репродуктивной активности принято считать расстояние между местом рождения и местом размножения для 95 % особей данного поколения. При исследовании перемещений многих животных было выяснено, что обычно около 95 % особей расселяются на расстояние меньшее, чем половина максимального (т. е. самого дальнего расселения). Разумеется, величины РРА у разных видов сильно варьируют. Ограничимся лишь небольшим рядом примеров:
дрозофилы (Drosophila sp.) разных видов – 150-400 м;
среднеазиатская черепаха (Testudo horsfieldi) –700 м;
зяблик (Fringilla coelebs) – 2 км;
черный коршун (Milvus migrans) – 1000 км;
крот (Talpa europaeus) – 44 м;
лисица (Vulpes vulpes) – 100 км;
песец (Alopex lagopus) – 850 км;
человек (сельские районы Европейской России, конец ХIХ в.) – 10 км.
У некоторых видов РРА подвержен колебаниям у особей разных поколений в разные периоды существования одной и той же популяции, в связи с чем имеют место временные флуктуации ареалов ряда видов, а также нерегулярные дальние миграции (естественные инвазии).
Лишь 5 % особей популяции выселяются за пределы ее РРА. Так называемые «особи-бродяги» иногда уходят очень далеко, порой на расстояния, на порядок превышающие величину РРА. Есть предположение, что склонность к «бродяжничеству» предопределена генетически; в таком случае это явление может иметь эволюционный смысл, обеспечивая потенциал расселения популяции, благодаря которому популяция может расширять свой ареал и осваивать новые территории.
2.3. Плотность популяции
Плотность – величина популяции, отнесенная к некоторой единице пространства; обычно выражается числом особей (реже биомассой) на единицу площади либо объема, например 500 деревьев/1 га, 4 зайца/1 км2, 200 экз. рыбы/1 га водоема и т. п.
Иногда различают среднюю плотность – численность на единицу всего учитываемого пространства – и экологическую, рассчитанную на единицу заселенного данным видом обитаемого пространства (биотопа). Например, если какой-то моллюск в водоеме заселяет только участки с илистым дном, то среднюю плотность придется рассчитывать для всей площади водоема, а экологическую – для суммарной площади заиленных участков.
Иногда на практике более значимым оказывается знание не конкретной плотности популяции в тот или иной момент времени, а наличие возможности оценить ее в сравнительном аспекте – близка она к максимальным показателям или минимальным, возрастает или уменьшается. В подобных случаях полезны показатели относительного обилия. Например, для оценки относительного обилия мышевидных грызунов в исследуемом биотопе выставляют в определенном порядке не менее 25 ловушек и выдерживают их в состоянии готовности не менее 3 полных суток, регулярно собирая «улов». Для сравнения полученных показателей общее число отловленных зверьков пересчитывают к стандартному показателю «число пойманных особей/100 ловушко-суток». Такие показатели по мере их накопления удобны для оценки динамики численности зверьков в разных биотопах, разные сезоны года и разные годы. Оценка абсолютной плотности по этой методике невозможна, и если в ней возникает необходимость, приходится прибегать к гораздо более сложным и трудоемким методам.
Приведенный пример является одним из множества методов оценки абсолютного и относительного многообразия организмов, определяющегося целым спектром особенностей как изучаемых видов организмов, так и биотопов, которые они заселяют.
Подробно рассматривать все существующие методы здесь не имеет смысла; их лучше постигать по мере возникновения надобности, пользуясь специальными методическими руководствами. В приведенном примере с грызунами для проведения исследования приходится прибегать к безвозвратному изъятию зверьков из биотопа; к счастью, это не является обязательным правилом, и большинство методов оценки плотности популяции основаны на подсчете организмов без их умерщвления.
2.4. Возрастная структура популяции
В популяциях большинства организмов можно обнаружить особей разного возраста. Возрастная структура популяции определяется соотношением в ней разных возрастных групп. Эта характеристика существенно влияет на темпы размножения и уровень смертности, а следовательно, на общую численность популяции и ее изменение во времени.
Под возрастной группой понимают группу особей одинакового астрономического либо физиологического возраста. Астрономический (календарный) возраст обычно исчисляют в годах (для недолго живущих организмов – в днях), для выражения физиологического возраста обычно определяют 3 возрастные группы: предрепродуктивную, репродуктивную и пострепродуктивную. При обсуждении различных аспектов возрастной структуры популяций часто используется также термин «поколение», под которым принято понимать непосредственное потомство особей, появившихся на свет на протяжении одного цикла размножения (у однократно размножающихся видов) или всего репродуктивного периода (у видов с неоднократным размножением на протяжении жизни). Впрочем, в отношении человека поколением нередко называют также группу одного астрономического возраста («поколение 2012 года рождения»).
Для выражения соотношения в популяции особей разного астрономического возраста используют так называемые возрастные (демографические) пирамиды (рис. 4; 5). По сути, это диаграммы, на которых число особей каждого возраста (или доля их в популяции) изображено полосой определенного масштаба. Полосы располагаются одна за другой в порядке увеличения значений возраста. Если есть необходимость представить соотношение возрастных групп раздельно по полу, полосы располагают горизонтально, одну над другой по мере увеличения возраста; при этом в левой части пирамиды принято располагать диаграмму для мужчин, в правой – для женщин (рис. 5).
Порой даже беглого взгляда на возрастную пирамиду бывает достаточно, чтобы составить представление об особенностях соотношения воспроизводства и смертности в данной популяции. В нормальной, устойчиво существующей популяции число молодых особей всегда больше, чем число особей старшего возраста; поэтому самой широкой частью такой пирамиды будет ее основание. Однако пирамида с широким основанием, но очень резко суживающаяся кверху (рис. 4, а), чаще всего характеризует популяции с очень высокой смертностью молодняка. Такая ситуация наблюдается, например, у всем известной большой синицы (Parus major): пара этих птиц за весну и начало лета обычно производит на свет два выводка общим числом около 20 потомков, однако до следующего сезона размножения в каждой семье доживают в среднем две птицы.
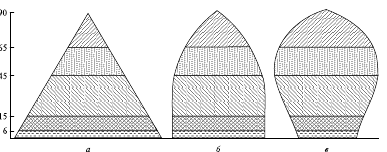
Рис. 4. Демографические пирамиды популяций, построенные без разделения особей по полу: а - растущая; б - стабильная; в - уменьшающаяся
Впрочем, популяция, оказавшаяся в особо благоприятных условиях и быстро растущая, зачастую тоже характеризуется высокой долей молодых особей. Однако по мере замедления темпов роста в такой популяции (с нарастанием ее плотности) либо снизится интенсивность размножения, либо возрастет смертность молодняка, и пирамида изменит свои контуры, приблизившись к варианту, изображенному на рис. 4, б. Однако существует очень мало популяций, у которых такой тип соотношения особей разного возраста в популяции сохранялся достаточно долго.
Есть данные, что близкое к подобному типу соотношение порой наблюдается у некоторых гидр (Hydrida), чья смертность в разном возрасте примерно одинакова. В популяции, численность которой снижается, будет содержаться большая доля старых особей (см. рис. 4, в). Похожий тип пирамид характеризует и некоторые сравнительно устойчивые популяции, если им свойственны высокая выживаемость молодых особей и смертность преимущественно в пострепродуктивном возрасте.
Возрастная структура популяции играет важную роль в демографических и социальных процессах в человеческом обществе. Она тесно взаимодействует с характеристиками воспроизводства человеческого населения. В принципе, все типы возрастных пирамид, отображенные на рис. 4, встречаются и у человека, характеризуя особенности воспроизводства, смертности и тенденции динамики этих характеристик в разных обществах и на разных этапах их развития.
Тип пирамиды, изображенный на рис. 4, а, характеризует страны с высокой рождаемостью и высокой смертностью. У возрастной пирамиды такого населения широкое основание (которое образует высокая доля детей в населении) и узкий шпилеобразный верх (малая доля доживающих до преклонного возраста), боковые стороны имеют вид вогнутых парабол, и чем сильнее вогнуты эти линии, тем выше уровень смертности в детском возрасте. Такой тип воспроизводства населения иногда называют примитивным (по многим параметрам, и не только демографическим, такое наименование ему вполне подходит). В далеком прошлом все народы имели этот тип воспроизводства населения, а некоторые страны сохраняют его и в настоящее время.
Социальное развитие общества приводит к снижению уровня смертности, а спустя 1-3 поколения – и рождаемости. На сравнительно небольшом отрезке времени возрастная структура общества приближается к типу, отраженному на рис. 4, б, а затем, с явным замедлением роста численности населения (о причинах которого пойдет речь ниже), она принимает форму колокола (см. рис. 4, в). Такой тип в демографии иногда называют неподвижным или стационарным.
Процесс снижения смертности рано или поздно замедляется или прекращается (смертность не может сокращаться бесконечно), в то время как снижение рождаемости будет продолжается. Начинается депопуляция — вымирание населения. Самая широкая часть возрастной пирамиды такой популяции постепенно все более смещается вверх, пирамида напоминает форму погребальной урны. Население стареет, т. е. в его составе увеличивается доля пожилых людей и сокращается доля молодежи. Такой тип воспроизводства населения в демографии называют регрессивным или деградационным.
В некоторых случаях возрастная пирамида популяции может иметь форму «елочки», когда длинные и короткие полосы чередуются (см. рис. 5).
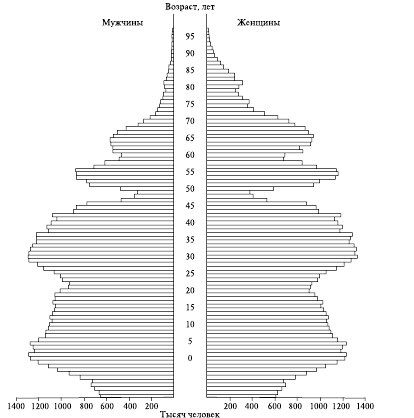
Рис. 5. Демографическая пирамида популяции, построенная с разделением особей по полу
Такой тип характерен для популяций тюленей в местах, где ведется интенсивный промысел недавно родившихся детенышей (так называемого белька), который проводится, как правило, один раз в 2-3 года, но при отсутствии должного контроля, во время которого часто происходит изъятие всего годового приплода.
Для определения физиологического возраста особей выделяют три категории (стадии):
1) предрепродуктивный возраст, когда молодые особи физиологически еще не способны к размножению (у серой крысы, например, до трех месяцев, у воробья до одного года, у гориллы до 5 лет, а у человека до 14 –16 лет);
2) репродуктивный (детородный) возраст, в котором особь способна к размножению (у человека до 45 –50 лет, в виде исключения дольше);
3) пострепродуктивный возраст, когда особь из-за старения организма уже неспособна к размножению.
У многих видов организмов стадия пострепродуктивного возраста отсутствует, например, вплоть до самой гибели сохраняют способность к размножению большинство деревьев, многие птицы. У многих насекомых, а также проходных лососевых рыб, размножение происходит один раз в жизни перед гибелью. В подобных случаях мы можем говорить об очень растянутом предрепродуктивном возрасте при отсутствии пострепродуктивного. У большинства же микроорганизмов выделение групп физиологического возраста на практике невозможно (в микробиологии иногда применяют понятие «возраст колонии микроорганизмов»).
У человека соотношение продолжительности стадий физиологического возраста в самой приближенной форме можно представить как 20:30:20 годам, т. е. в пропорции 1:1,5:1, тогда как у млекопитающих (по крайней мере в условиях неволи) эта пропорция обычно равна 1:2:3. Такое сопоставление породило предположение о потенциальном среднем возрасте человека 120 лет, который не реализуется лишь из-за воздействия неблагоприятных внешних и внутренних факторов.
В популяционной экологии растений принят несколько иной подход к определению возрастной структуры. В составе ценопопуляций выделяют четыре возрастных периода:
1) латентный (семена);
2) прегенеративный (всходы и т. д.);
3) генеративный (размножающиеся растения);
4) постгенеративный (неразмножающие, вплоть до отмирающих).
Ценопопуляции, включающие растения всех возрастных периодов, называются нормальными; они способны к полному самоподдержанию и не зависят от внешнего поступления семян. Популяции же, состоящие преимущественно из особей прегенеративного периода, сильно зависят от заноса зачатков извне. Они характерны для начальных этапов освоения пространства в биоценозе и носят название инвазионных.
Кажущиеся принципиальными различия между особенностями возрастной структуры популяций у растений и животных на самом деле не столь резки, во время исследования колорадского жука Leptinotarsa decemlineata было обнаружено, что часть особей в некоторых популяциях может впадать в состояние затяжной, многолетней диапаузы, а за этот период в активно развивающейся популяции сменится не один десяток поколений. Затем «спящие» жуки по каким-то причинам активизируются и быстро восстанавливают популяционную численность (в том случае, если она, например, была сокращена химическими методами защиты).
2.5. Половая структура популяций
По отношению к животным, подавляющее большинство которых раздельнополы, а также к двудомным растениям (ива, облепиха, тополь и др.), применимо понятие половой структуры популяции, которая определяется численным соотношением самцов и самок в разных возрастных группах. Различают первичное, вторичное и третичное соотношение полов.
Первичное соотношение полов определяется соотношением половых хромосом в процессе мейоза. Обычно наблюдается соотношение 1:1, но нередки и отклонения, которые могут быть обусловлены спецификой определения пола у тех или иных групп организмов, а также полиплоидией и влиянием на половые хромосомы аутосом.
У человека и многих других млекопитающих пол определяется наличием или отсутствием Y-хромосомы. Особи с хромосомными аномалиями (XXY, X0 и др.) в популяциях встречаются редко и соотношение 1:1 нарушают незначительно. Но, например, в популяциях леммингов кроме нормальных ХХ-самок существует некоторое количество ХY-особей, также функционирующих как самки, поскольку локализованный в аутосомах фактор подавляет действие Y-хромосомы. Схожая картина наблюдается у некоторых рыб, например меченосцев.
У многих беспозвоночных (в частности, у Drosophila melanogaster) наличие Y-хромосомы само по себе не определяет пол. Пол этих организмов в основном зависит от численного соотношения определенных хромосом, и отклонения в первичном соотношении полов здесь могут быть гораздо более значительными.
Явление полиплоидии также играет роль фактора, заметно влияющего на первичное соотношение полов. Например, в ряде популяций серебряного карася (Carassius auratus) преобладают триплоидные самки (3n = 160 – 166), а в некоторых регионах (в том числе в Беларуси) самцы у этого вида вообще отсутствуют, и популяции размножаются партеногенетически.
Вторичное соотношение полов — соотношение полов на момент рождения либо ко времени завершения метаморфоза. Отличия вторичного соотношения полов по сравнению с первичным зависят, как правило, от действия факторов внешней среды. У некоторых видов определяющими являются абиотические факторы. Например, у многих амфибий температура воды в период развития икры влияет на соотношение полов у головастиков. А у морского кольчатого червя Bonellia viridis определяющими являются гомотипический биотический фактор: его личинка, если в период свободной жизни ей удастся прикрепиться к взрослой самке, развивается в самца, если же этого не происходит, она оседает на дно и формируется как самка. Другой интересный пример демонстрирует кольчатый червь Ophiriotrocha puerilis, который меняет пол в течение жизни. Молодые особи сначала являются самцами, а когда количество сегментов достигает 15-20, они превращаются в самок.
Иногда вторичное соотношение полов регулируется гормонально. Так, при развитии раздельнополых близнецов у коровы рождаются самец и бесплодная телка-интерсекс, так как мужские гормоны развивающегося самца подавляют развитие половых органов у самки.
Третичное соотношение полов определяется числом самцов и самок среди половозрелых, размножающихся особей, а также особей пострепродуктивного возраста. Отклонения от соотношения 1:1 здесь нередко очень значительны и обычно определяются неодинаковой смертностью самцов и самок в процессе взросления. Поскольку показатели смертности могут существенно не совпадать даже в разных популяциях одного вида, соответствующие различия фиксируются и в третичном соотношении полов.
Для человека и многих других млекопитающих, а также птиц, характерна общая закономерность: уменьшение доли самцов в старших возрастных группах, что обычно связано с более высокой смертностью мужских особей. Эта закономерность хорошо иллюстрируется на примере человеческого населения нашей страны.
Иногда наблюдаются весьма необычные ситуации с третичным соотношением полов. Например, в некоторых популяциях горностая (Mustela erminea) самцы спариваются с еще слепыми, недавно родившимися самками, к этому времени имеющими зрелые яйцеклетки. После такого оплодотворения, хотя и с некоторой задержкой, происходит развитие эмбрионов и рождение вполне нормальных детенышей.
2.6. Этологическая структура популяций
По отношению к животным, характеризующимся более-менее выраженными особенностями поведения, применимо подразделение популяции по признакам поведения входящих в нее особей, т. е. этологической структуры.
Этологические группировки могут выделяться по различным признакам. Наиболее простым является разделение особей на группы по особенностям двигательной активности. Такие группы легко выделяются, например, у ряда видов насекомых, в популяциях которых имеются как крылатые, способные расселяться на большие расстояния формы, так и бескрылые (либо с редуцированными крыльями) особи.
Для ряда высших животных характерно образование более или менее устойчивых «семейных» ячеек из самцов, самок и молодняка (семейные стаи у волков, прайды у львов, стада и табуны у копытных и т. д.). Подобные образования принято объединять под обобщенным названием демы. Поведение особей внутри демов различается: им присущи определенные отношения иерархии, разделение функций между особями и т. п., что вызывает и некоторые особенности в поведении, легко заметные даже при простейших наблюдениях за такими группами. При этом у многих видов наряду с «семейными группами» встречаются и особи, ведущие преимущественно одиночный образ жизни (старые самцы у копытных, неполовозрелые особи у хищных птиц и др.) или же однополые группы не размножающихся особей (группы молодых самцов у морских котиков, стаи неполовозрелых птиц у журавлей и др.). Специфику такой структурированности популяции отражает ее разделение по репродуктивной агрегированности и образу жизни.
С данным подразделением тесно связано деление популяции по поведению внутри групп. Этот подход акцентирует внимание не на характере семейных связей внутри групп, а на особенностях поведения конкретных особей. Исследуя популяцию в таких аспектах, удается выявлять особенности иерархии особей внутри группы, проявления агрессивного поведения, которое может обеспечивать охрану индивидуального участка, влияет на подбор полевых партнеров и ситуации, связанные с конкурентными отношениями за пищу. Нередко при таком подходе удается обнаружить и определенные формы поведенческой специализации отдельных особей внутри группы, например по характеру и способам поиска корма, по тактике избегания хищника и т.п.
Широко известным является разделение популяции по системам брачных отношений. Ключевыми признаками для выявления таких структур является характер брачных отношений и спариваний внутри популяции. Известны следующие системы брачных отношений:
1) моногамия: брачную пару создают один самец и одна самка. При этом пара может образоваться либо на короткий период спаривания (лягушки, многие рыбы, кряква, леопард), либо на один полный цикл размножения (пингвины, аисты, дикие гуси, большинство воробьиных птиц), либо на более длительный период, обычно до гибели одного из партнеров (лебеди, лисы, волки). Гибель партнера в последнем случае не исключает из процесса размножения оставшуюся особь пары, и через какое-то время она находит нового партнера;
2) полигамия: одна особь вступает в брачную связь с двумя и более особями противоположного пола. Различают несколько типов полигамии:
полигиния, когда один самец вступает в брачные связи с несколькими (иногда многими) самками. Видимо, наиболее часто встречающийся тип полигамии у птиц и млекопитающих (африканский страус, крапивник, морские котики, олени и др.);
полиандрия, когда одна самка имеет брачные связи с несколькими самцами. Нередко встречается у рептилий (ящерицы, змеи) и очень редко – у теплокровных животных. Из числа последних наиболее известен пример обитающего в северных тундрах кулика-плавунчика (Phalaropus lobatus), его самка, образовав пару с самцом, спаривается с ним и в построенное за короткий срок гнездо откладывает четыре яйца, после чего покидает своего партнера и ищет нового, с которым спаривание, строительство гнезда и откладка яиц повторяются. Насиживание кладки и вождение выводка целиком ложится на самца. Интересно, что, в отличие от других птиц, самки у этих куликов окрашены более ярко, чем самцы;
полибрахигиния, когда каждый самец спаривается с несколькими самками, а каждая самка – с несколькими самцами. Типичная полибрахигиния наблюдается у птиц-полигамов с хорошо выраженным ритуалом токования (глухарь и тетерев, турухтаны), кукушки, бродячих домашних собак;
промискуитет – структура брачных отношений, при которых любая особь имеет равную вероятность скрещивания с любой другой особью противоположного пола. Типичным примером являются многощетинковые черви и иглокожие, выпускающие свои гаметы просто в море. Определенно сходный тип (но безотносительно к полу) наблюдается и у ветроопыляемых растений. Обычно эти системы брачных отношений причисляют к видовым характеристикам, но в природе они часто сочетаются в одних и тех же популяциях. Например, у многих птиц (выпь, крапивник, дроздовидная камышевка) наряду с самцами-полигамами, на участках которых гнездится по несколько самок, наблюдаются и самцы моногамы, и даже территориальные самцы, остающиеся холостыми. В таких случаях говорят о факультативной полигамии.
Нередко в популяции при детальном наблюдении за входящими в нее особями встречаются определенные предпочтения, связанные с выбором полового партнера.
В таких случаях можно говорить о структуре скрещиваний внутри популяции. Сравнительно редко структура скрещиваний носит случайный характер (панмиксия). Примером панмиктического скрещивания являются иглокожие, выбрасывающие свои половые продукты прямо в воду. Чаще всего наблюдаются так называемые неслучайные структуры скрещиваний.
По генетическому характеру выделяют инбридинг (гомогамию) – скрещивание большей частью генетически схожих особей, и аутбридинг (гетерогамию) — скрещивание генетически различных организмов. Если виду присущ выраженный полиморфизм по каким-либо хорошо заметным внешним признакам, проявления гомогамии или гетерогамии порой можно наблюдать непосредственно в природе. Например, в по
Дата добавления: 2022-04-12; просмотров: 151;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
