Наследственный материал клетки. Хромосомы и уровни её организации.
Наследственный материал клетки- это, то благодаря чему мы отличаемся друг от друга, то что дается нам от родителей, а им от их и так далее. Огромный и наверняка претерпевший сильные изменения за большой промежуток времени он передался вам. Наследственный материал содержится в наших ДНК, заглядывая наперед скажу, что ДНК безусловно важная структура, но без белоксинтезирующего аппарата(информационная РНК, рибосомы, транспортная РНК) он бы не значил для нас ничего, так как ваше телосложение похожее скажем на отца, а голубой цвет глаз матери – это белки- результат экспрессии (проявляемости) гена. Ген- это участок молекулы ДНК, которая содержит информацию о структуре, кодируемого белка, который в организме будет служить либо ферментом( например, гидролитические ферменты лизосомы), либо регулятором какого- то процесса жизненного процесса (регулятор митоза, репликации и.т.д), либо будет частью клетки(структурный белок, например, белок плазматической мембраны). ДНК в свою очередь расположено в хромосомах и занимают примерно 50-55% её объема, остальное приходится на структурные гистоновые(щелочные) и негистоновые (кислые) белки, которые укорачивают цепь ДНК. Наверняка задается вопрос: «Зачем это нужно? Какой смысл?» . Если бы не было подобной упаковки, то ДНК попросту не поместилась в клетке, ведь её истинная длина около 2 м. , вследствие чего выделяют 6 уровней организаций:
1) Нуклеосомный уровень- это, когда 8 гистоновых белков(Н2А, Н2В, Н3, Н4 по две штуки каждая) образуют шар, по- научному белковый кор. И вокруг этого кора наматывается ДНК ,делая вокруг неё 1,75 оборота , благодаря чему он укорачивается в 2-4 раза.
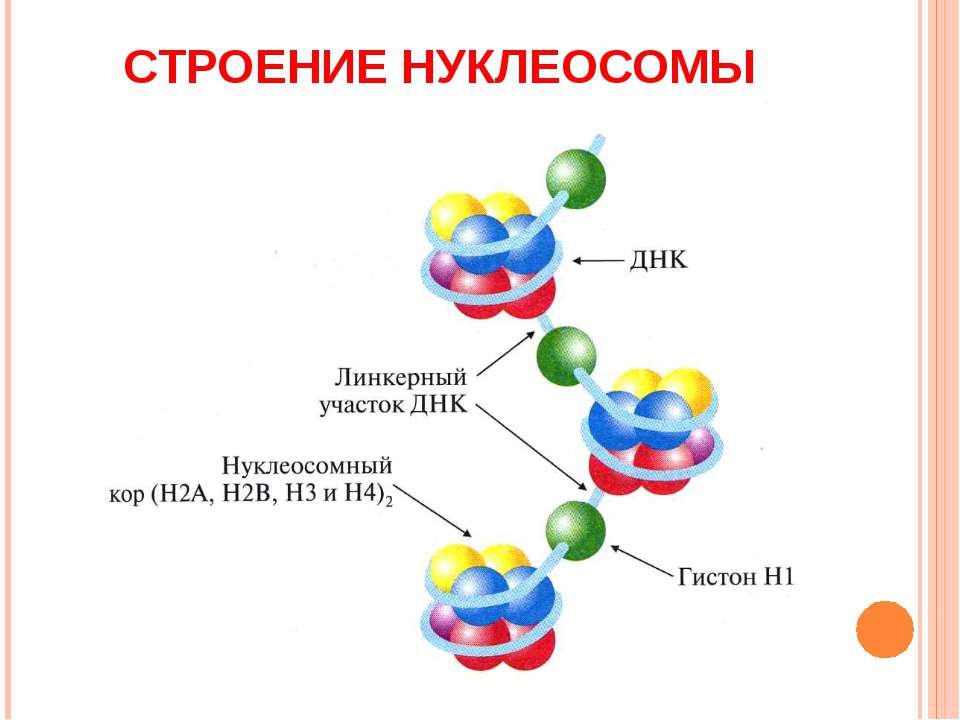
2) Нуклеомерный уровень организации. На картинке вы видите линкерные учаастки (участки между корами). В данном уровне организации они занимают ключевое место, так там будет находится еще один гистоновый белок Н1, который будет притягивать друг к другу эти коры и тем самым укорачивать ещё сильнее длину ДНК в 6-7 раз. Получившуюся в итоге структуру также называют хроматиновой фибриллой или соленоидным хроматином.
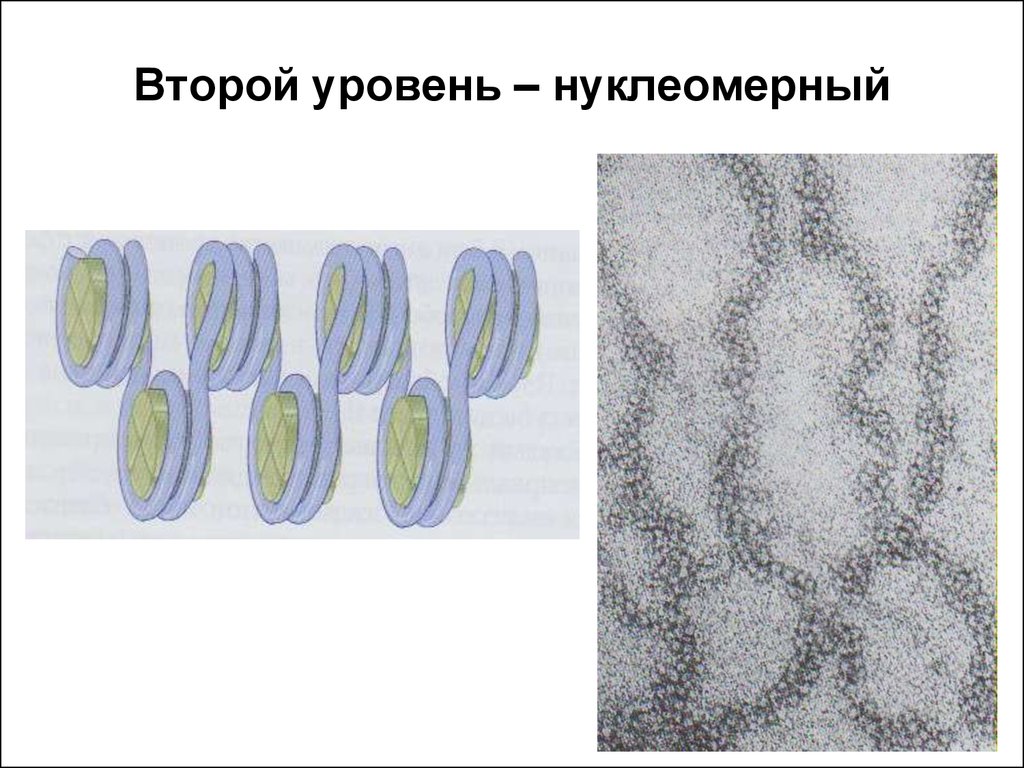
3) Следующий уровень укорочения- хромомерный уровень организации. На данном уровне укорочения уже участвуют негистоновые (кислые) белки. Укорочение на этом уровне достигается образованием петлевых структур.
4) Хромонемный уровень организации- дальнейшее укорочение ДНК достигается благодаря сближению петель и образование петлевых доменов. Этот уровень можно увидеть в интерфазе- период до деления клетки.
5) Хроматидный уровень организации-дальнейшая петлизация хромонемы и образование хроматиды.
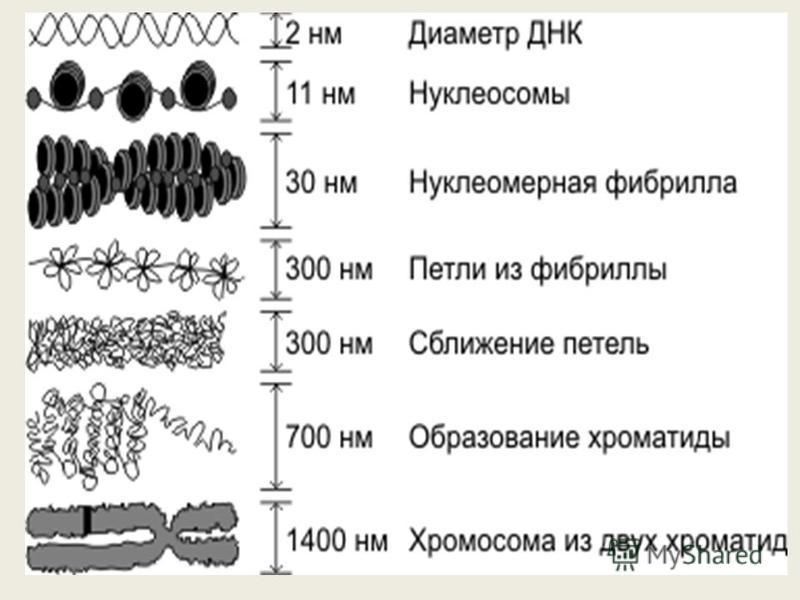
6) Хромосомный и самый высший уровень организаций. Хромосома- структура, состоящая из 2 хроматид, соединенных по середине центромерой. Концевые участки хромосомы называются теломерами. Теломеры заняты гетерохроматином – плотный хроматин, с которого не осуществляются процессы транскрипции, как и центромерные участки. Промежуток между этими двумя частями(между центромерой и теломерой) занято эухроматином- с которого осуществляются процессы транскрипции, потом трансляции и синтезируется белок, следовательно ген проявляется.
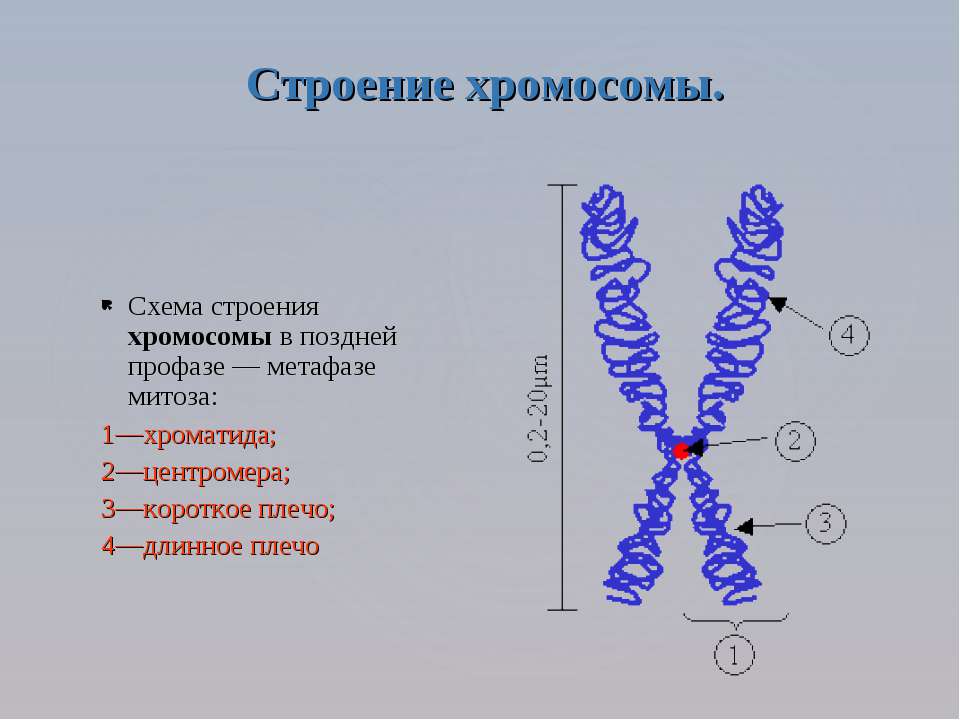
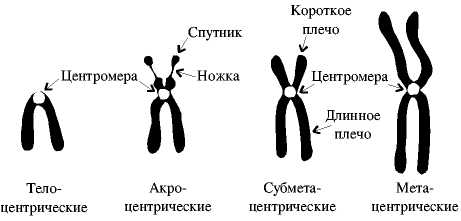
В зависимости от положения центромеры хромосомы подразделяются на:
1) Метацентрические хромосомы- центромера располагается по середине, вследствие чего плечи(смотри на картинку) равной длины.
2) Субметацентрические- плечи немного отличаются в длине. В такой хромосоме уже различают какое плечо длинное, а какое короткое.
3) Акроцентрические- плечи сильно различаются в длине.
4) Телоцентрические- всего 1 плечо, центромера на краю. Подобных хромосом в организме как правило нет. Их наличие- приводит к патологии.
Хромосомы можно изучать благодаря остановке митоза на стадии метафазы, когда они таковыми представлены(хорошо видимы) и изучить на наличие хромосомных мутации.
Понятие о гене. Строение гена эукариот и прокариот, их сравнительная характеристика.
Что мы называем геном и где они расположены? Гены в количестве примерно 30000 располагаются в ДНК, те в хромосомах, те в ядре. История формирования первичных представлении о гене берет начало с работ Менделя, которые к тому времени по его предположению носили название «наследственных факторов», отвечающих за развитие конкретного признака. Позже эти «наследственные факторы» В.Иогансеном в 1909 г. получили название гены. Работы Менделя стали основой для формирования классических представлений о структуре гена и формирования постулата: «Один ген- один признак». В соответствии с представлениями классической генетики, ген- неделимая структурно- функциональная единица наследственности, отвечающая за развитие какого-то признака(цвет глаз например). Но, эти представления в дальнейшем были опровержены экспериментами Серебровского и Дубинина, Им были показаны, что при облучении, что при облучении мух рентгеновскими лучами возникали различные мутации в пределах одного гена, отвечающего за щетинки на теле мух. У одной группы при облучении наблюдался усиленный рост щетинок, у второй- укорочение, у третьей- отсутствие и четвертой- сохранились в норме. В соответствии с представлениями классической генетики Менделя ожидалось увидеть только третий вариант- отсутствие щетинок из- за мутации этого гена. Эти данные привели к пониманию сложности строения гена и формированию нового постулата (формулировка) «Один ген- одна полипептидная цепь(белок)». Ген не всегда определяет признак, порой для этого нужно несколько, а не один, белков.
Строение гена. Ген- участок молекулы ДНК, состоящий из 2 частей: регуляторного и кодирующего участков. Разделены они между собой последовательностью в 300 нуклеотид, называемой точкой начала транскрипции. По отношению к этой точке разделяют +(правее- кодирующая часть) и – (левее- регуляторная часть) последовательности. Функция регуляторной части- контролирует начало и интенсивность процесса транскрипции- образовании информационной РНК(иРНК)- матрица(как бы то, на что опираются, прототип) для синтеза белка. Кодирующая часть- матрица для синтеза иРНК. Прошу пожалуйста ради собственного блага смотреть картинки, которые я вставляю, так как это поможет вам при освоении материала.
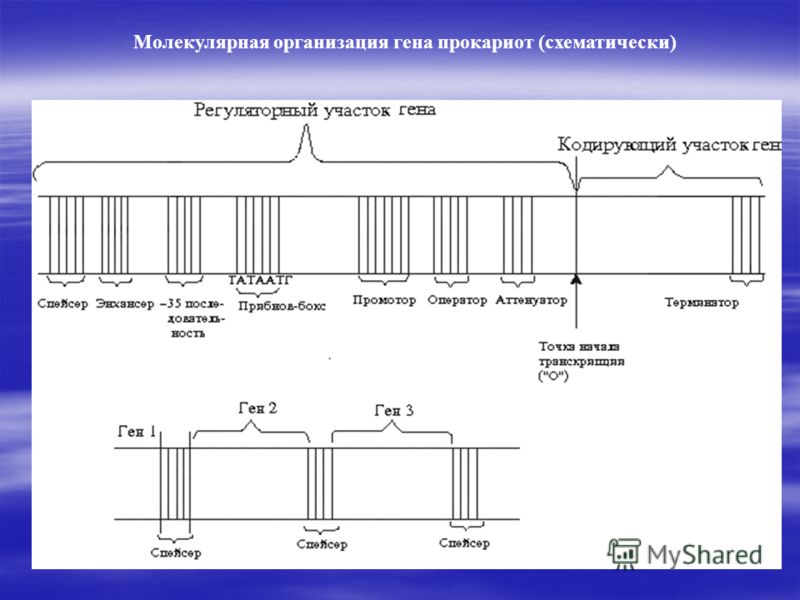
Итак, регуляторная часть у прокариот содержит следующие нуклеотидные последовательности:
1) Промотор- последовательность нуклеотидов, с которым связывается РНК- полимераза- белок, осуществляющий синтез иРНК и состоящее из 2 частей:
а) Прибнов- бокс(-10 последовательность(-10 значит, что она находится на расстоянии в 10 нуклеотидов от точки начала транскрипции )) – с ним связывается РНК- полимераза.
б) узнаваемая последовательность( -35 последовательность)- с ним связывается сигма субъединица РНК- полимеразы, которая является как бы активатором процесса транскрипции;
2) оператор- последовательность нуклеотидов, с которыми связываются регуляторные белки. Если в этом участке имеется регуляторный белок, то процесса транскрипции не происходит;
3) аттенуатор- последовательность нуклеотидов, запускающая или предотвращающая процесс транскрипции
4) энхансер- последовательность нуклеотидов, увеличивающая скорость транскрипции
5) сайленсер (на картинке его нету, но знайте, что он распологается ближе энхансера )- последовательность нуклеотидов, снижающая скорость транскрипции.
Кодирующая часть гена прокариот состоит только из экзонов- информативных кодируемых участков и терминатора- последовательность нуклеотидов(АТТ, АТЦ, АЦТ), служащая сигналом об окончании транскрипции, когда РНК- полимераза доходит до сюда, он дальше не идет.
Строение гена эукариот. Строение гена эукариот мало чем отличается от прокариот, в ней имеются в основном все те же последовательности нуклеотидов за некоторыми исключениями. Так у эукариот нету оператора, а промотор состоит из 3 частей:
1) ТАТА- бокс(Голдберга- Хогнесса бокс, -20 последовательность)- с ней связывается РНК-полимераза;
2) ЦААТ- бокс(-60 последовательность)
3) ГЦ-бокс(- 80 последовательность)
ЦААТ и ГЦ- боксы регулируют скорость транскрипции.
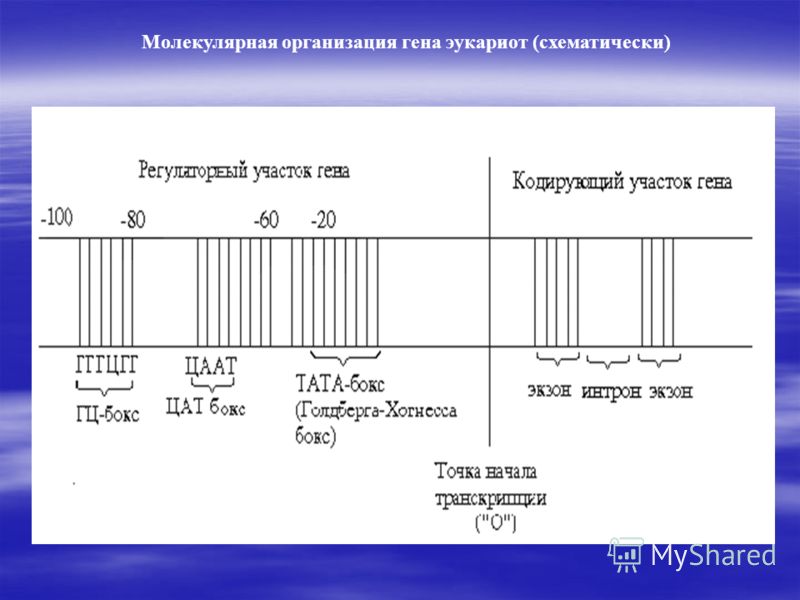
Кодирующая часть эукариот состоит еще из интронов - некодируемых последовательностей, но не спешите с выводами, так как они выполняют 2 важные функции:
1) Они снижают частоту мутации;
2) Обуславливают процесс альтернативного сплайсинга (об этом позже, не зациклывайтесь )
В человеческом организме 30000 генов, но они занимают всего 2 % хромосом, так что гены разделены между собой обширными нетранскибируемыми нуклеотидными последовательносями- спейсарами.


Теперь, когда вы знаете строение гена я бы хотел вернуться к опыту Серебровского и Дубинина (прочитайте ещё раз, если забыли). Чрезмерный рост щетинок был обусловлен мутацией в энхансере , медленный рост- мутацией в сайленсере, отсутствие – в промоторе, а когда изменений не происходило в интроне, так как эта часть не кодируется(не участвует в определений структуры кодируемого с гена белка). Если вы поняли предпоследнее предложение, значит хорошо усвоили эту тему.
Репликация (удвоение молекулы ДНК). Значение теломер (концевые участки хромосом)
Репликация- это сложный процесс самоудвоения молекулы ДНК. В неделящейся клетке наследственный материал можно выразить 2n2c(n-количество хромосом, а с- количество ДНК). В интерфазе происходит процесс репликации, следовательно наследственный материал становится 2n4c. Данный процесс протекает по определенным нерушимым правилам, или как принято говорить принципам репликации:
1) Комплементарность- это когда аденину одной цепи противопоставлен тимин другой (между ними 2 водородные связи), гуанину- цитозин(между ними 3 водородные связи);
2) Полуконсервативность- когда в результате деления одна цепь старая(материнская цепь), а вторая- новая (дочерняя цепь) ;
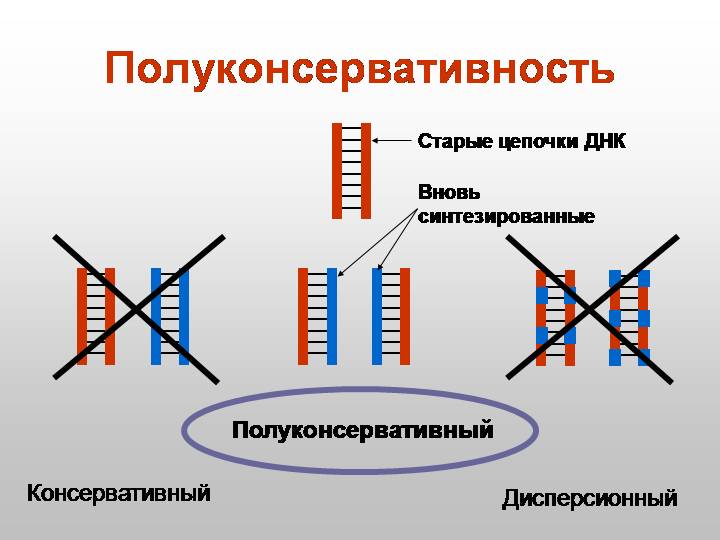
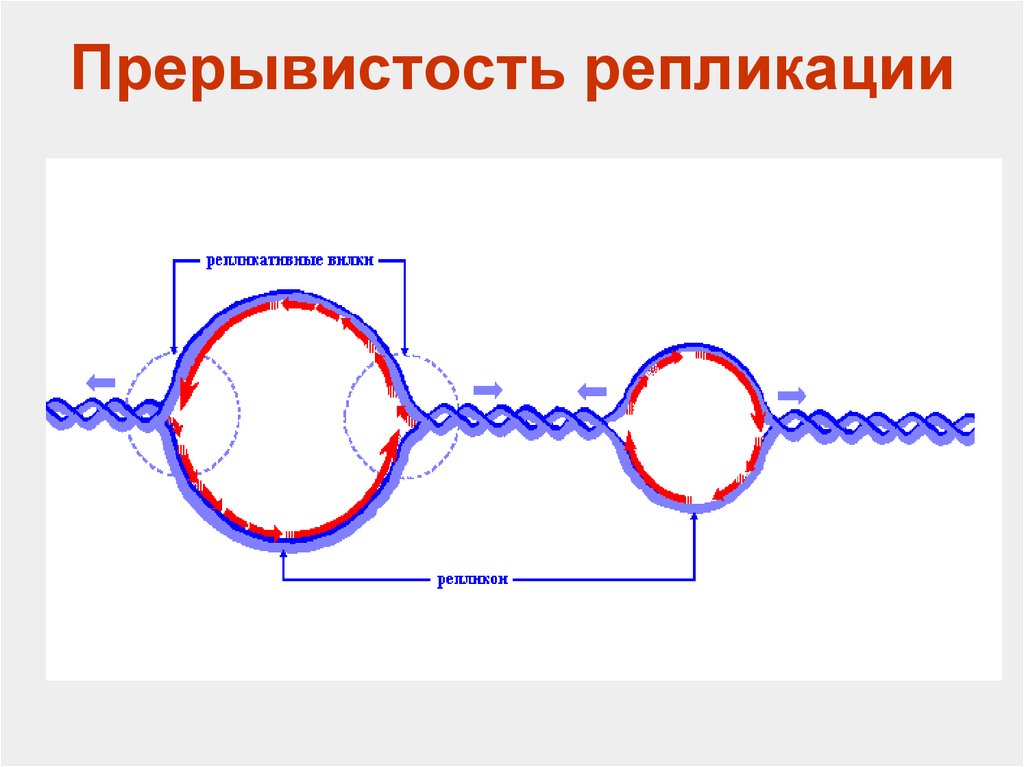
3) Прерывистость- ДНК настолько обширна, что если бы репликация начиналась на одном месте и заканчивалась в другом, то это бы заняло очень большое количество времени, вследствие чего начинается в нескольких местах.
Для вашего понимания 4 и 5 принципов прежде я должен вам напомнить строение нуклеотида- «кирпичики», из которых состоит ДНК.
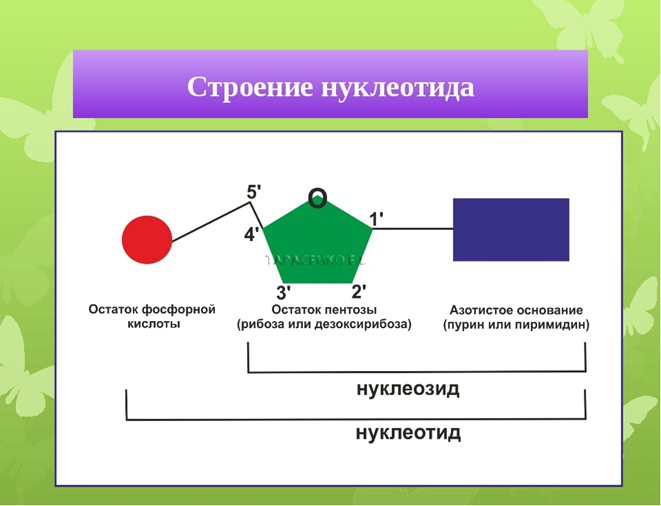
Нуклеотид состоит из азотистого основания (А,Т,Ц,Г), пятиуглеродного сахара- дезоксирибозы и остатка фосфорной кислоты. Когда изображают ДНК в схематическом виде вы увидите 5̕ и 3̕. Что они значат? Немного химии: на 3̕ имелась группа ОН, но она соединилась с другой такой же группой ОН, но не дезоксирибозы , а фосфорной кислоты, далее выделяется Н20 и образуется прочная ковалентная связь, это значит что удлинение молекулы ДНК происходит с 5̕ → 3̕ конец.
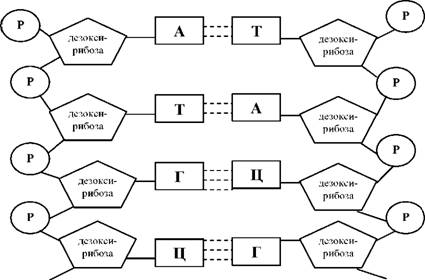
51---АТТГАТАЦАГГЦ---31
31 ---ТААЦТАТГТЦЦГ---5̕
(Схематическое изображение ДНК)
4) Итак, следующий принцип- антипараллельность- это если, одна цепь с 5̕ -3̕, то вторая будет с 3̕ - 5̕ (смотри выше)
5) Униполярность- синтез дочерних цепей осуществляется всегда с 5̕ -3̕ .
Процесс репликации идет в 3 стадии:
1) Инициаиция
2) Элонгация
3) Терминация
Теперь перейдем к самому процессу. Так как репликация идет прерывисто образующиеся в результате структуры называются репликонами, а зоны репликации- репликационными вилками. Предшествует образованию репликативной вилки образование прежде репликационного глазка в области точек начала репликации. При этом от точки начала репликации могут образоваться либо 1 репликационная вилка(у эукариот) или 2 у прокариот. Образование репликативного глазка и есть 1 этап- инициация репликации.
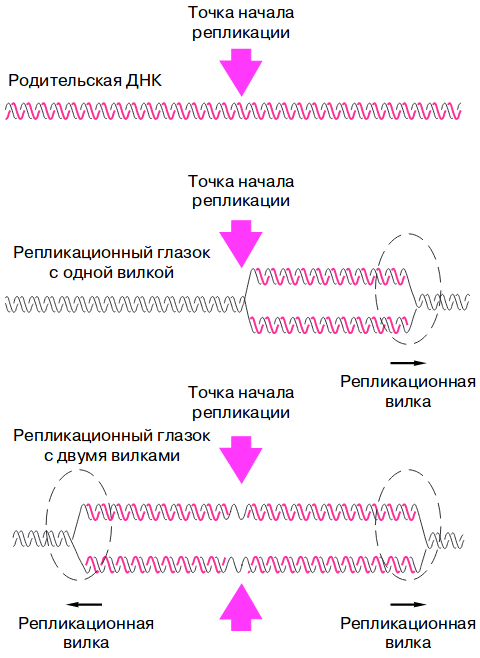
Элонгация- основной этап процесса репликации. Она осуществляется благодаря целому каскаду ферментов, без которых этот процесс невозможно было бы представить. При репликации одна дочерняя цепь синтезируется непрерывно, потому что её направление совпадает с направление хеликазы, а 2 дочерняя- кусками- фрагментами Оказаки. Синтезирующаяся непрерывно цепь называется лидирующей(2 на рисунке), а фрагментами- отстающей(1 на рисунке).
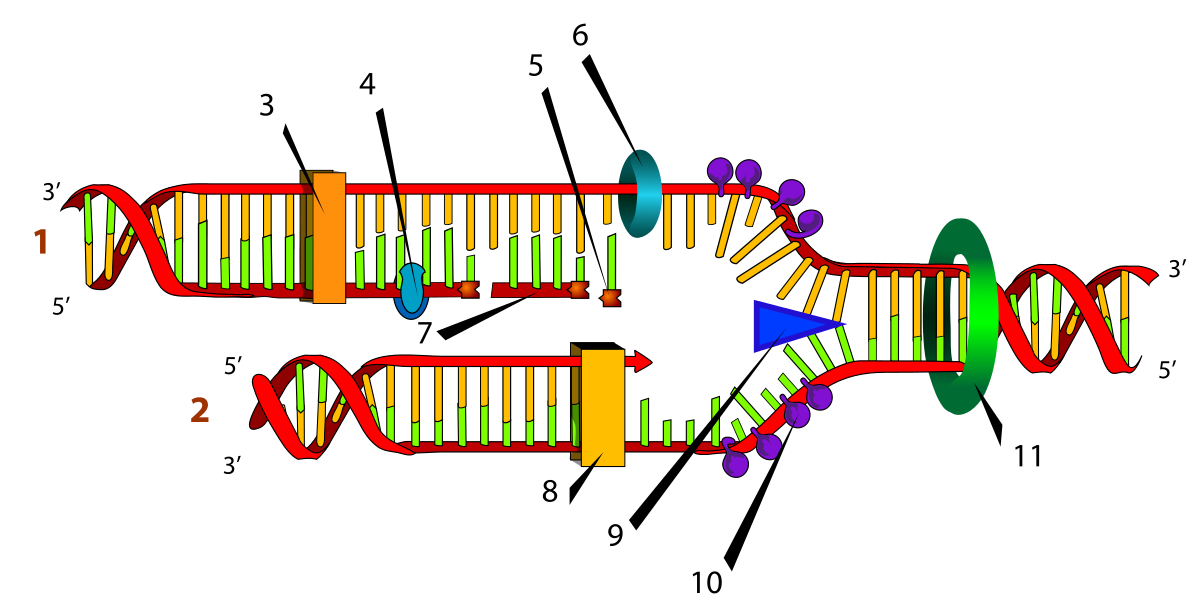
1) Хеликаза- функцией этого фермента является расплетение двух нитей ДНК, на рисунке она обозачена под цифрой 9.
2) Топоизомераза- когда хеликаза разрывает две нити, то в них накапливается напряжение и они начинают быстрее вращаться- явление суперспирализации. Топоизомераза специально предназначена ради того, чтобы её снять. На рисунке под цифрой 11.
3) ДНК-полимераза- синтезирует дочернюю цепь(на рисунке под цифрами 3 и 8)
4) ДНК- лигаза – сшивает фрагменты Оказаки.
5) РНК- праймаза- синтезирует РНК- затравки(РНК- праймеры), которые выступают в роли маяка для ДНК-полимераз, указывая им откуда начать, и только после присоединения к затравке ДНК- полимераза может начать синтез дочерней цепи. При этом для лидирующей цепи нужно всего один праймер, а дочерней- множество.(6 на рисунке)
6) SSB- белки- нужны для того, чтобы удерживать разъединившиеся цепи вдали друг от друга, чтобы те не соединились вновь.(10 на рисунке)
По некоторым данным в процессе репликации принимают участие около 20 белков, но здесь перечислены основные.
Терминация репликации. В ДНК имеются специфические последовательности, с которыми связываются терминаторные белки, препятствующие дальнейшему продвижению репликативной вилки. Синтез ДНК заканчивается.
Значение теломер для репликации. Как оказалось ДНК-полимераза начинает синтез только со специального РНК-праймера. После окончания синтеза ДНК РНК-праймеры удаляются, а пропуски в одной из дочерних цепей ДНК заполняются ДНК-полимеразой. Однако на 3'-конце ДНК такой пропуск заполнен быть не может, и поэтому 3'-концевые участки ДНК остаются однотяжевыми, а их 5'-концевые участки – недореплицированными, вследствие чего одна из 2 дочерних цепей становится короче на 60-80 нуклеотидов.
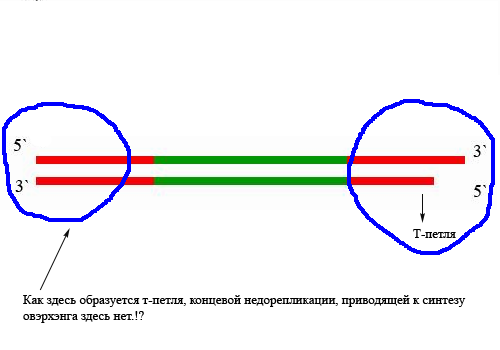
Так происходит практически у всех клеток живого организма, за исключением опухолевых, половых и стволовых клеток(в костном мозгу- синтезируют клетки крови) за счет наличия специального фермента, называемого теломеразой. Его работа состоит из 2 этапов: элонгации и транслокации. Элонгация – удлинение теломерного участка, а транслокация- его перемещения для следующей элонгации.
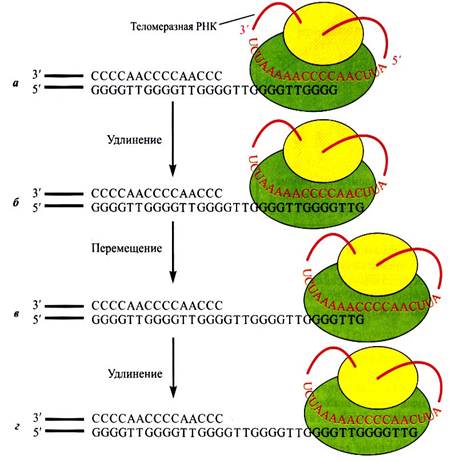
Благодаря их наличию эти клетки никогда не стареют. Существует даже теломерная теория старения, которая утверждает , что жизнь клетки прямо пропорционально длине теломер. Теломеры- это плотно упакованный нетранскибируемый гетерохроматин(красный ), который защищает ценный эухроматин(фиолетовый).

Так что существует определенное число делений после которого теломера исчезает и клетка погибает. Это число называется лимитом Хейфлика.
Транскрипция(синтез и созревание иРНК)
Транскрипция- сложный процесс синтеза иРНК на матрице одной из 2 цепей ДНК. Транскрипция наряду с трансляцией являются механизмами экспрессии( проявляемость, свидетельство активности) генов и в результате этой экспрессии синтезируется белок- продукт активности гена. Но о трансляции позже, уверен, если вы дочитали до сюда, то начальное представление вы о ней уже имеете. Вернемся к транскрипции, как и у репликации есть нерушимые правила(принципы) транскрипции:
1) Комплементаность- у иРНК тоже есть те же азотистые основания, как и у ДНК, за исключением тимина, поэтому аденину- урацил, цитозину- гуанин.
2) Консервативность- иРНК одноцепочечная и эта цепь, так как одна не содержит старой цепи как при репликации, поэтому он называется принципом консервативности, а не полуконсервативности
3) Антипараллельность- это если, одна цепь с 5̕ -3̕, то вторая будет с 3̕ - 5̕ .
4) Униполярность- синтез иРНК всегда происходит в направлении 5̕ -3̕ .
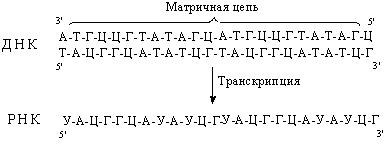
Не запутайтесь матрицей для синтеза иРНК на рисунке послужила 1 цепь(3 и 4 принципы).
Процесс транскрипции осуществляется в 3 этапа:
1) Инициация
2) Элонгация
3) Терминация
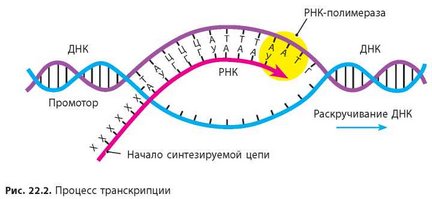
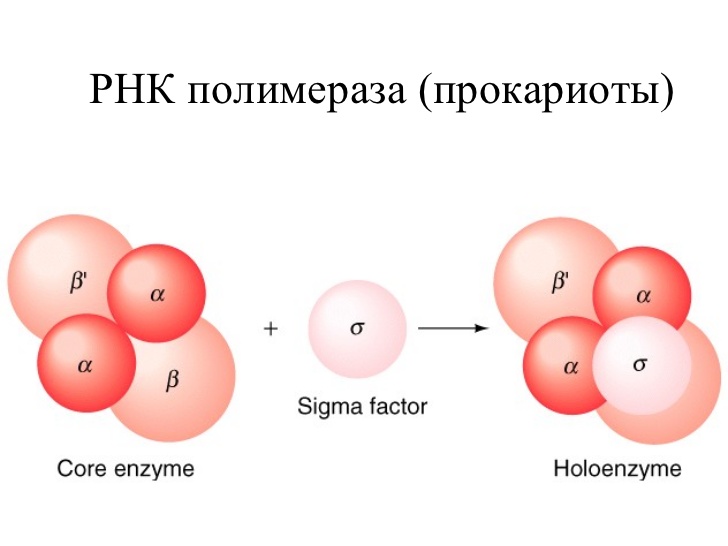
Основным ферментом синтеза иРНК является фермент РНК- полимераза, которая состоит из 2 альфа и 2 бета субъединиц. Также в ней особое место занимает сигма субъединица, которая активирует этот комплекс и как видите на рисунке кор-фермент(пассивный) становится холо-ферментом(активный)
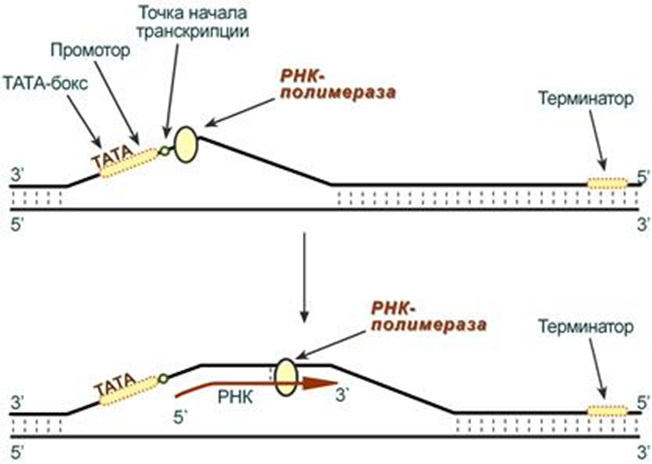
Для начала процесса инициации необходимо, чтобы образовался холо-фермент на промоторе. При этом вначале присоединяется кор- фермент к ТАТА- боксу(- 10 последовательность) и лишь после сигма субъединица к узнаваемой последовательности(-35 последовательность) после чего между двумя цепями начинают разрываться водородные связи и ДНК начинает раскручиваться как на рисунке. У эукариот процесс инициации протекает несколько сложнее, так как для образования этого комплекса им нужен целый ряд ферментов- называемых для удобства факторами транскрипции. И если у прокариот имеется всего 1 разновидность РНК-полимеразы, синтезирующая все 3 типа РНК(тРНК, иРНК, рРНК), то у эукариот их 3. Потом пройдете, что за что отвечает. И если вспромнить строение гена, то на процесс транскрипции как и у прокариот, так и у эукариот могут влиять на процессивность: энхансеры(усиливают процесс), сайленсеры(тормозят процесс).
Элонгация. Эта стадия основная. Холо- фермент продвигается в направлении кодирующей части ДНК и после синтеза 8-10 первых нуклеотидов иРНК на матрице 1 цепи ДНК сигма субъединица отрывается и дальнейший синтез иРНК осуществляет кор-фермент. Свою функция активатора сигма субъединица выполнила и теперь может идти отдыхать. На данном этапе, как и при инициации у эукариот, как и у прокариот, так и эукариот существуют факторы элонгации, которые ускоряют этот процесс.
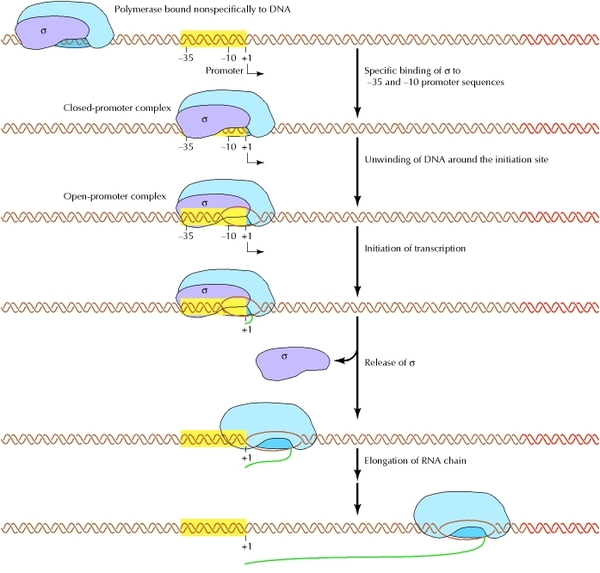
Терминация транскрипции. Окончание процесса транскрипции происходит тогда, когда РНК- полимераза синтезирует один из трех терминирующих кодона(УАА, УГА, УАГ).
Дата добавления: 2022-07-20; просмотров: 175;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
