ЕДИНЫЙ ПРОЦЕСС ИЛИ РАЗЛИЧНЫЕ СОСТОЯНИЯ?
В 1953 г. Е. Азеринский и Н. Клейтман открыли феномен «быстрого» сна, и тем самым – новую эру в изучении сна. Если раньше пассивные и активные теории регуляции сна рассматривали бодрствование как состояние, противоположное сну, а сон сам по себе считался единым феноменом, то теперь идея монолитного сна оказалась разрушенной и стали изучаться механизмы как медленного, так и быстрого сна. В итоге в настоящее время регуляторные процессы медленного сна связываются со структурами промежуточного мозга, а быстрого сна – главным образом со стволовыми структурами моста.
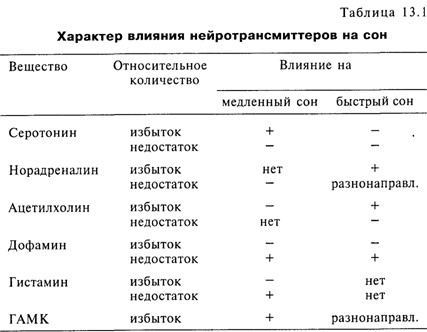
В 60–70 гг. М. Жуве, основываясь на обширных исследованиях с пересечениями и повреждениями мозга, а также фармакологических и нейроанатомических данных, предложил моноаминергическую теорию регуляции цикла «сон–бодрствование», согласно которой медленный и быстрый сон связаны с активностью различных групп моноаминергических нейронов – в регуляцию медленного сна включены серотонинергические нейроны комплекса шва, в то время как норадренергические нейроны ответственны за наступление быстрого сна. Впоследствии было показано участие различных нейротрансмиттеров в регуляции медленного и быстрого сна. В табл. 13.1 представлены эти данные [Drucker-Colin, Merchant-Nancy, 1995].
Различие механизмов медленного и быстрого сна подтверждается также и в нейрогуморальных концепциях сна, основоположником которых является А. Пьерон. Ещё в начале текущего столетия, на основании результатов своих экспериментов на собаках, у которых сон вызывался введением спинномозговой жидкости других собак, лишённых сна в течение нескольких суток, А. Пьерон предположил, что наступление сна связано с накоплением в организме определённых веществ (гипнотоксинов). Впоследствии «фактор сна» многочисленные исследователи выделяли из спинномозговой жидкости, крови и мочи различных животных, и с каждым годом увеличивался список обнаруженных в организме веществ, связанных со сном. В табл. 13.2 представлены все пептиды, изучавшиеся на предмет влияния на сон. Р. Друкер-Колин и Н. Мерчант-Нэнси [Drucker-Colin, Merchant-Nancy, 1995], суммировав полученные данные, объясняют обилие этих веществ тем, что все они действуют через посредство какого-то ещё неизвестного механизма, ответственного за наступление сна, а единственного фактора сна в понимании А. Пьерона реально не существует.
Ко всем перечисленным веществам нужно добавить мелатонин, который выделяется эпифизом только ночью и также играет важную роль в поддержании сна (о механизме действия различных групп веществ на сон см. обзор [Lemoine, Allain, 1996]).
Таким образом, результаты обширных нейрофизиологических, нейрохимических и нейрогуморальных исследований свидетельствуют не только о сложности и многообразии взаимодействия различных факторов в регуляции цикла «сон–бодрствование», но и о различии механизмов медленного и быстрого сна.

Дата добавления: 2016-06-05; просмотров: 2153;
Поиск по сайту
Узнать еще
- AMD Opteron (Socket 940), Intel Xeon Dempsey и Xeon Woodcrest (Socket 771): популярные серверные двуядерные процессоры.
- Base-5 или толстый Ethenet
- CIH — газораспределительный механизм с распределительным валом в головке блока цилиндров
- Cемейство Мотыльковые (Fabaceae, или Leguminosae)
- Cравнительная характеристика усилителей на БТ
- D-технология построения чертежа. Типовые объемные тела: призма, цилиндр, конус, сфера, тор, клин. Построение тел выдавливанием и вращением. Разрезы, сечения.
- I-s диаграмма рабочего процесса ГТД
- I. Диалектический характер процесса познания

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
