И передающиеся трансмиссивно
В эту экологическую группу паразитических простейших входит большое количество видов из классов Жгутиковые и Споровики, имеющих сложные циклы развития. Все переносчики этих паразитов являются специфическими. Распространенность заболеваний приурочена к зонам встречаемости переносчиков. Некоторые паразиты этой группы поселяются только в организме человека, поэтому соответствующие заболевания являются антропонозными. Другие формируют устойчивые природные очаги, поражая разные виды диких животных. Цист эти паразиты не образуют и непосредственно во внешнюю среду не выделяются. Диагностика заболеваний, вызываемых этими паразитами, основана на обнаружении их трофозоитов в крови и биоптатах тканей или на иммунологических реакциях.
Профилактика включает общие меры профилактики трансмиссивных заболеваний (борьба с переносчиками) и конкретные, зависящие от видовой специфики паразитов.
Лейшмании Leischmania (кл. Жгутиковые) — возбудители лейшманиозов. Заболевания человека вызываются несколькими видами и подвидами паразитов, которые объединяются в четыре комплекса: L. donovani — возбудитель висцерального лейшманиоза, L. tropica — возбудитель кожного лейшманиоза, L. mexicana — возбудитель лейшманиоза Центральной Америки, L. brasiliensis — возбудитель бразильского лейшманиоза. Все виды сходны морфологически и имеют одинаковые циклы развития. Они существуют в двух формах: в безжгутиковой, или лейшманиальной, и жгутиковой; или промастиготной (рис. 19.8).
Лейшманиальная форма очень мелка — 3—5 мкм в диаметре. Характерной чертой ее является круглое ядро, занимающее около '/4 цитоплазмы; жгутика нет, но перпендикулярно клеточной поверхности располагается палочковидный кинетопласт. Эти формы обитают в клетках ретикулоэндотелиальной системы человека и ряда млекопитающих (грызунов, собак, лис). Промастиготная форма удлинена — до 25 мкм, спереди находится жгутик, у основания которого хорошо виден такой же кинетопласт, что и в безжгутиковой стадии паразита. Обитает в пищеварительной системе москитов. Безжгутиковая форма, посеянная на культуральную среду, превращается в жгутиковую.
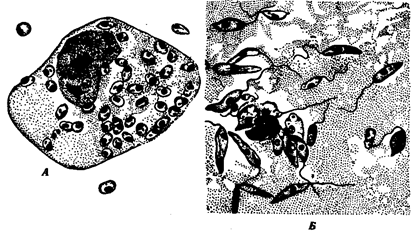
Рис. 19.8. Лейшмания. Безжгутиковая (А) и жгутиковая (Б) формы
Лейшманиозы широко распространены в странах с тропическим и субтропическим климатом на всех континентах там, где обитают москиты. Они—типичные природно-очаговые заболевания (см. § 18.13). Природными резервуарами являются грызуны, дикие и домашние хищники. Заражение человека происходит при укусе инвазированными москитами.
По патогенному действию лейшманий заболевания, которые они вызывают, делят на три основные формы: кожный, слизисто-кожный и висцеральный лейшманиозы.
При кожном лейшманиозе очаги поражения находятся в коже. Это самый распространенный тип лейшманиоза, протекающий относительно доброкачественно. Возбудителями кожного лейшманиоза в Африке и Азии являются L. tropica, а в Западном полушарии — L. mexicana и ряд штаммов L. brasiliensis. Лейшманий L. tropica и L. mexicana вызывают на коже длительно не заживающие язвы на месте укусов москитами. Язвы заживают через несколько месяцев после образования, а на их месте на коже остаются глубокие рубцы. Некоторые формы L. brasiliensis способны распространяться по лимфатическим сосудам кожи с образованием многочисленных кожных язв в отдалении от мест укусов.
Слизисто-кожный лейшманиоз вызывается подвидом L. brasiliensis brasiliensis. При этой форме заболевания паразиты проникают из кожи по кровеносным сосудам в носоглотку, гортань, мягкое нёбо, половые органы, поселяются в макрофагах соединительных тканей этих органов и вызывают здесь деструктивные воспаления.
Висцеральный лейшманиоз вызывает L. donovani. Заболевание начинается через несколько месяцев или даже лет после заражения как системная инфекция. Паразиты размножаются в макрофагах и в моноцитах крови. Нарушаются функции печени, кроветворение. Очень велика интоксикация. При отсутствии лечения заболевание заканчивается смертью.
Лабораторная диагностика основана на микроскопировании мазков из кожных язв при кожном и слизисто-кожном лейшманиозах, пунктатов лимфатических узлов и костного мозга при висцеральном лейшманиозе. В окрашенных препаратах обнаруживается лейшманиальная форма паразитов как внутри клеток, так и внеклеточно. В сомнительных случаях производят посев материала, взятого от больного, на специальную культуральную среду, на которой лейшманий приобретают промастиготную форму, активно передвигаются и легко обнаруживаются при микроскопировании. Используют также и биологические пробы — заражение лабораторных грызунов.
Профилактика — в первую очередь, это борьба с переносчиками и уничтожение природных резервуаров (грызунов и бродячих собак), а также профилактические прививки.
Trypanosoma brucei gambiense и T.b. rhodesiense (кл. Жгутиковые) — возбудители африканского трипаносомоза, или сонной болезни. Паразит имеет извилистую заостренную с обеих сторон форму. Длина его 17—28 мкм. Стадии, паразитирующие у человека, имеют один жгутик, ундулирующую мембрану сбоку и хорошо заметный кинетопласт у основания жгутика.
Трипаносомы поселяются у человека в крови, лимфе, спинномозговой жидкости, в тканях головного и спинного мозга и в серозных полостях. T.b. gambiense встречается в Западной Африке, а T.b. rhodesiense — в Восточной и Юго-Восточной Африке.
Жизненный цикл этих паразитов протекает в организме человека, домашних и диких млекопитающих, в первую очередь копытных. T.b. gambiense чаще поражает человека, свиней и собак, T.b. rhodesiense — диких животных — антилоп и носорогов. Переносчиком первого подвида является муха це-це ,Glossina palpalis, живущая поблизости от жилища человека, второго— G. morsitans, обитающая в открытых саваннах и саванновых лесах. В связи с этим сонная болезнь, возбудителем которой является T.b. gambiense, встречается в антропогенных очагах культурных ландшафтов. Ежегодно регистрируется около 10 000 новых случаев заражения. Восточноафриканский трипаносомоз распространен значительно реже в естественной природе. В основном заболевают охотники, туристы, сезонные рабочие, каждый год — около 1500 человек.
Сонная болезнь без лечения протекает около 5 лет и выражается в нарастающей мышечной слабости, депрессии, истощении и сонливости. Возможны случаи самоизлечения, но обычно заболевание заканчивается смертью больного.
Восточноафриканский трипаносомоз протекает более злокачественно, длится не более 6 мес. и также заканчивается смертью.
Для паразитирования трипаносом у млекопитающих и человека характерны циклические подъемы интенсивности инвазии за счет их размножения, сопровождающиеся изменениями строения и антигенных свойств паразитов. Во время увеличения количества паразитов в крови преобладают трипаносомы удлиненной формы. Антигены, которые они образуют, вызывают формирование антител в организме хозяина. Под действием антител многие паразиты гибнут и интенсивность инвазии снижается. Выжившие трипаносомы укорачиваются и начинают вырабатывать другие антигены. Укороченные формы паразита, инвазионные для мухи це-це, в ее организме вновь приобретают удлиненную форму, инвазионную для человека. Изменение формы тела и смена антигенных свойств оболочки повторяются многократно. Таким образом, популяция паразита в хозяине выживает и избегает его иммунной реакции.
Антигенные свойства поверхности трипаносомы зависят только от одного белка — гликопротеина, полностью покрывающего всю клетку. Гликопротеин построен из 470 остатков аминокислот. Каждая новая волна размножения паразитов представляет собой новую популяцию трипаносом, обладающих новым поверхностным антигеном. Эти вариации антигенных свойств помогают паразиту преодолевать иммунный ответ хозяина и делают невозможной вакцинацию населения, обитающего в природных очагах трипаносомозов.
Смена антигенных свойств обеспечивается заменой поверхностных гликопротеинов, кодируемых разными генами, относящимися к одному мультигенному семейству. Один клон трипаносом может образовывать попеременно до 100 разных варьирующих гликопротеинов. Гены этих протеинов возникли в процессе эволюции, вероятно, путем дупликаций и последующей дифференцировки, как и другие семейства генов. В геноме трипаносомы имеется сайт экспрессии, в который поочередно перемещаются гены поверхностных гликопротеинов, приближаясь к промотору, обеспечивающему их специфическую активацию. Там они транслируются. Не исключено, однако, что сайт экспрессии в геноме трипаносомы не единственный, и даже возможно, что разные гены гликопротеинов активируются несколькими механизмами. В любом случае речь идет о своеобразной адаптации паразита к специфическим условиям существования, повышающей его выживаемость и открывающей ему широкие эволюционные перспективы.
Лабораторная диагностика — исследование мазков крови и спинномозговой жидкости больного для выявления в них возбудителя. Используются также иммунологические реакции и заражение лабораторных животных.
Профилактика — кроме борьбы с переносчиками применяют профилактическое лечение здоровых людей, живущих в очагах трипаносомоза, делающее организм невосприимчивым к инвазии. Практиковавшийся ранее отстрел диких животных, являющихся природным резервуаром паразита, вряд ли рационален в связи с возможным нарушением экологического баланса, который складывался в биогеоценозах на протяжении тысячелетий.
Trypanosoma cruy — возбудитель американского трипаносомоза, или болезни Чагаса. Длина этой трипаносомы в крови человека достигает 20 мкм. Кинетопласт очень крупный, округлой формы. Характерной особенностью возбудителя является способность к внутриклеточному паразитизму. При этом трипаносомы проникают вначале в макрофаги кожи и слизистых оболочек, а затем и в клетки миокарда, нейроглии и мышц, теряя жгутики, ундулирующие мембраны, и превращаясь в безжгутиковые, или амастиготные, формы. Здесь и происходит размножение паразитов. В крови эти трипаносомы никогда не делятся. В конечном счете, пораженная клетка вся заполняется амастиготными формами трипаносом и разрывается, а паразиты инвазируют новые клетки. При этом часть их, превращаясь вновь в жгутиковую форму, поступает в кровь, откуда в дальнейшем они могут попасть в организм переносчика (рис. 19.9).
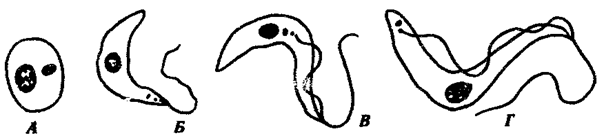
Рис. 19.9. Трипаносома. Безжгутиковая (А) и жгутиковые {Б—Г) формы
Переносчиками являются триатомовые клопы pp. Triatoma, Rhodnius и Panstrongylus (см. разд. 21.2.2). В них трипаносомы размножаются и достигают состояния инвазионности, поступая в заднюю кишку. Вскоре после кровососания клопы испражняются на покровы человека или животного и трипаносомы проникают в кровь через раневое отверстие от хоботка или через неповрежденные слизистые оболочки губ, носа и глаз. Окончательными хозяевами кроме человека являются броненосцы, опоссумы, крысы, обезьяны и домашние животные — собаки, кошки и свиньи.
Болезнь поражает в основном детей младшего возраста, у которых протекает остро. В старшем возрасте заболевание переходит в хроническую форму.
Патогенное действие выражается в поражении органов, в клетках которых развиваются паразиты: характерны миокардиты, кровоизлияния в мозговые оболочки и менингоэнцефалит. Иногда заболевание протекает легко и заканчивается самопроизвольным излечением.
Диагностика — в острой форме заболевания возможно обнаружение трипаносом в крови. При хроническом течении рационально введение крови больного морским свинкам, у которых они обнаруживаются в большом количестве на 14-е сутки. Существует еще и своеобразный метод диагностики — кормление на больном неинвазированных переносчиков-клопов, в кишечнике которых трипаносомы быстро размножаются и легко обнаруживаются. Применяют также и методы иммунодиагностики.
Трипаносомы представляют большой интерес не только потому, что способны вызывать у человека серьезные, смертельно опасные заболевания. Колоссальная экологическая пластичность, обеспечивающая им эффективное паразитирование в организме хозяина на протяжении нескольких лет, в условиях постоянно действующих механизмов иммунитета, позволяет этим паразитам осваивать и новых хозяев. Так, описанная в последние годы вспышка трипаносомоза в Эфиопии была вызвана адаптацией к человеку Т.b. brucei — подвида, паразитирующего обычно только у крупного рогатого скота и антилоп. Та же причина, вероятно, лежит в основе существования вспышек трипаносомозов в Индии и Малайзии, где у больных были выделены паразиты, обитающие обычно в крови грызунов и низших обезьян. В Центральной и Южной Америке кроме Т. cruzy известна и еще одна трипаносома — Т. rangeli, которая чаще паразитирует у кошек и собак, но способна инвазировать и человека, вызывая у него нетяжелую, быстро проходящую лихорадочную реакцию.
Малярийные плазмодии Plasmodium (кл. Споровики) — возбудители малярии. Известны следующие виды малярийных плазмодиев, паразитирующие у человека: Р. vivax — возбудитель трехдневной малярии, Р. falciparum — возбудитель тропической малярии, Р. malariae — возбудитель четырехдневной малярии, Р. ovale— возбудитель овале-малярии, близкой к трехдневной. Три первых вида широко распространены в тропических и субтропических климатических поясах, последний — только в тропической Африке. Все виды сходны морфологически и жизненными циклами, отличаясь друг от друга деталями строения и некоторыми особенностями цикла развития, проявляющимися в основном продолжительностью его отдельных периодов.
Жизненный цикл малярийных плазмодиев типичен для споровиков, включая стадии бесполого размножения в виде шизогонии, полового процесса и спорогонии. Окончательным хозяином паразитов является комар р. Anopheles (см. разд. 21.22), а промежуточным — только человек. Комар является одновременно и переносчиком. Поэтому малярия — типичное антропонозное трансмиссивное заболевание.
Со слюной зараженного комара при укусе плазмодии попадают в кровь человека (рис. 19.10). Развитие паразитов в организме человека происходит синхронно. С током крови они разносятся по организму и поселяются в клетках печени. Здесь они растут и размножаются шизогонией таким образом, что один паразит делится на тысячи дочерних особей. Клетки печени при этом разрушаются и паразиты, называющиеся на этой стадии мерозоитами, поступают в кровь и внедряются в эритроциты. С этого момента начинается эритроцитарная часть цикла развития плазмодия. Паразит питается гемоглобином, растет и размножается шизогонией. При этом каждый плазмодий делится на 8—24 мерозоита. После разрушения эритроцита мерозоиты попадают в плазму крови и оттуда в новые эритроциты, после чего весь цикл эритроцитарной шизогонии повторяется.
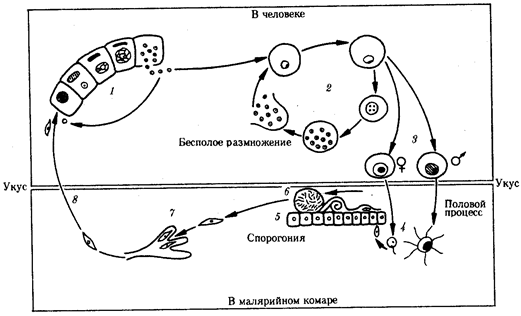
Рис. 19.10. Жизненный цикл малярийного плазмодия:
1 — преэритроцитарная шизогония в клетках печени, 2 — эритроцитарная шизогония, 3 — образование гаметоцитов, 4 — оплодотворение, 5 — спорогония в стенке желудка комара, 6 — овоциста со споромитами, 7 — проникновение спорозоита в слюнные железы комара, 8— заражение человека
Из части мерозоитов в эритроцитах образуются незрелые половые клетки — мужские и женские гаметоциты. Они являются инвазионной стадией для комара. Дальнейшее их развитие возможно только в его пищеварительной системе. При укусе больного человека комаром гаметоциты попадают в желудок последнего, где из них образуются зрелые гаметы. В результате оплодотворения в желудке комара образуется подвижная зигота, которая перемещается на наружную поверхность стенки желудка и покрывается оболочкой, формируя ооцисту. С этого момента начинается период спорогонии, когда содержимое ооцисты многократно делится, образуя около 10000 спорозоитов — тонких серповидных клеток, которые после разрыва оболочки поступают в слюнные железы комара. При кровососании спорозоиты поступают в кровяное русло человека.
Таким образом, в организме человека плазмодий размножается только бесполым путем — шизогонией, человек является его промежуточным хозяином. В организме комара проходят две другие стадии цикла развития паразита: половой процесс — гаметогония и образование спорозоитов за счет деления под оболочкой ооцисты — спорогония. Поэтому малярийный комар является окончательным хозяином этого паразита.
Выход большого количества мерозоитов из эритроцитов сопровождается выбросом в плазму крови значительной массы токсических продуктов жизнедеятельности. Их воздействие на организм приводит к резкому повышению температуры, ознобу, слабости и головным болям. Такое состояние возникает внезапно и длится в среднем 1,5—2 ч. Вслед за этим наступает чувство жара, сухость во рту, жажда. Температура тела достигает 40—41°С. Через несколько часов все перечисленные симптомы исчезают, и больные обычно засыпают. Весь приступ может продолжаться от 6 до 12 ч. При трехдневной и овале-малярии промежутки между приступами составляют 48 ч, число таких приступов может достигать 10—15, после чего они прекращаются за счет повышения уровня специфического иммунитета, но паразиты в крови еще могут обнаруживаться. В таком случае человек становится паразитоносителем и продолжает представлять опасность для окружающих как возможный источник заражения.
Естественный отбор приводит к возникновению новых антигенных вариантов возбудителя, которые обеспечивают возможность наступления рецидивов заболевания. Рецидивы могут повторяться несколько раз, но постепенно популяция эритроцитарных паразитов полностью погибает. Однако в течение 3—5 лет инвазия может вновь активизироваться за счет находящихся в латентном состоянии в печени экзоэритроцитарных шизонтов, которые могут выходить из печеночных клеток и внедряться в эритроциты. Таким образом, весь процесс болезни может начаться снова.
При малярии, вызываемой Р. malariae, приступы повторяются через 72 ч. Часто встречается и бессимптомное носительство. Экзоэритро-цитарной стадии в цикле развития этого паразита нет, поэтому поздние рецидивы невозможны, хотя инвазия характеризуется упорным течением и длится до 40 лет.
При тропической малярии вначале приступы развиваются через разные промежутки времени, а позже — через 24 ч. От осложнений со стороны центральной нервной системы или почек возможна смерть больного. Шизонты в клетках печени не сохраняются, а заболевание может продолжаться до 18 мес.
Все виды малярийных плазмодиев могут инвазировать человека и при гемотрансфузии (переливание крови). В этом случае ни у одного из паразитов не формируется экзоэритроцитарной стадии. Поэтому поздних рецидивов в этом случае не бывает. Гемотрансфузионный способ заражения наиболее часто встречается при четырехдневной малярии в связи с тем, что при этой форме болезни шизонты в эритроцитах находятся в очень малом количестве и могут не обнаруживаться при исследовании крови доноров.
Иногда человек может быть инвазирован одновременно двумя или тремя видами плазмодиев. В таком случае малярийные приступы не имеют четкой периодичности и клинический диагноз затруднен.
Лабораторный диагноз малярии можно поставить только в период, соответствующий стадии эритроцитарной шизогонии, когда в крови удается обнаружить паразитов.
Плазмодий, недавно проникший в эритроцит, имеет кольцевидную форму. Его цитоплазма выглядит как ободок, окружающий крупную вакуоль с продуктами диссимиляции. Ядро паразита смещено к краю клетки. Следующая стадия называется амебовидным шизонтом. У паразита появляются ложноножки, а вакуоль увеличивается. Наконец плазмодий занимает почти весь эритроцит. Следующая стадия развития паразита — фрагментация шизонта. На фоне деформированного эритроцита обнаруживаются множественные мерозоиты, в каждом из которых лежит ядро. Кроме бесполых клеток в эритроцитах можно увидеть и гаметоциты. Они отличаются крупными размерами, не имеют псевдоподий и вакуолей (рис. 19.11).
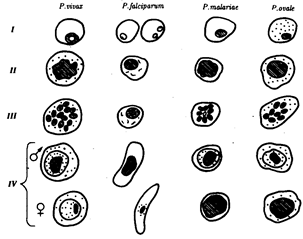
Рис. 19.11. Малярийные плазмодии. Стадии развития в эритроцитах:
I—стадия кольца, II—стадия амебовидного шизонта, III—стадия фрагментации, IV—гаметоциты
Профилактика малярии — раннее выявление и лечение больных, профилактическое лечение в зонах широкого распространения малярии. Как и при любых трансмиссивных заболеваниях, необходима прицельная борьба с переносчиками.
Дата добавления: 2016-08-23; просмотров: 1945;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
