ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ЖИВЫХ ОРГАНИЗМОВ
Комплекс экологических факторов, составляющий специфику любого местообитания, требует от населяющих это место организмов комплекс специфических приспособлений. Поэтому в процессе исторического развития животные и растения приобрели специфические черты, затрагивающие особенности строения, обмена веществ, динамику жизненных процессов и т. п. Все эти особенности определяют внешний облик организмов. В природе можно часто видеть, как разные виды приспосабливаются к сходным условиям среды. Такие типы приспособления выражаются в определенном морфологическом строении организмов и называются жизненными формами. Среди приспособлений организмов к условиям среды, возникших в результате эволюции, наиболее наглядными можно считать адаптации, проявляющиеся в особенностях внешнего строения растений и животных. Их называют морфологическими (от греч. morphē – форма). Определенные типы внешнего строения, возникшие как приспособления к экологическими условиям местообитаний, называют жизненными формами организмов.
Жизненные формы растений и животных очень разнообразны. Они выделяются по совокупности признаков строения и образа жизни. Так, наиболее широко распространенные жизненные формы растений – деревья, кустарники, травы. Последние делятся на водные и наземные, среди которых, в свою очередь, также выделяются разнообразные формы. Яркие примеры приспособлений к суровым условиям среды дают такие жизненные формы растений, как суккуленты (в засушливом климате), лианы (при недостатке света), стланики и растения-подушки (в тундрах, высокогорьях с низкой температурой и сухостью при сильных ветрах).
Жизненные формы животных выделяются по разным признакам для разных систематических групп. Так, для зверей одними из основных признаков для выделения жизненных форм, помимо среды обитания, считаются способы передвижения (ходьба, бег, прыжки, плавание, ползание). Характерными чертами внешнего строения наземных прыгунов, например, являются длинные задние конечности с сильно развитой мускулатурой бедер, длинный хвост, короткая шея. К ним обычно относятся обитатели открытых пространств: азиатские тушканчики, австралийские кенгуру, африканские прыгунчики и другие прыгающие млекопитающие, живущие на разных континентах.
Жизненные формы птиц различают по типу их местообитания и способу добывания пищи, а рыб – в основном по форме тела. Жизненные формы обитателей водоемов также выделяют по типу их местообитаний. В водной толще мелкие организмы образуют планктон (от греч. planktos – блуждающий), т. е. совокупность организмов, живущих во взвешенном состоянии и неспособных противостоять течениям. Обитатели грунта образуют бентос (от греч. bentos – глубина). К отдельным жизненным формам относятся организмы, живущие у поверхностной пленки воды или на различных твердых субстратах.
Похожие жизненные формы возникли в результате эволюции, происходящей в сходных экологических условиях у систематически разных организмов: например, кенгуру и тушканчики, дельфины и рыбы, птицы и летучие мыши, черви и змеи и т. д.
Нельзя считать, что, претерпев ряд глубоких изменений в процессе эволюции и достигнув большого разнообразия, живая природа застыла в неизменном облике. Она продолжает меняться. И эта способность организмов к изменению является важнейшим фактором, обеспечивающим соответствие между организмами и средой их обитания.
6. Температура как экологический фактор
Из всех климатических факторов, связанных с энергетикой биосферы, наибольшее экологическое значение имеет температура. Она результирует преобразования потока энергии у поверхности Земли и, в свою очередь, оказывает существенное влияние на энергетику биоты. Температура воздуха на Земле колеблется в диапазоне от -88,3 (ст. «Восток», Антарктида) до +58,7 °С (Гарьян, Ливия). Температура поверхности песка или камня в пустыне может достигать +70 °С. Средняя годовая температура приземного слоя воздуха над континентами и океаном (исключая Антарктиду) – +15,7 °С.
Средняя расчетная температура самой биомассы суши принимается равной +17,5 °С. В области восточно-сибирского антициклона амплитуда сезонных изменений температуры достигает 100 °С, а в зоне экваториальных дождевых лесов – всего 2–4 °С. В почве температурные колебания заметно сглажены: на глубине 25 см суточный ход температуры уже отсутствует. Еще более стабильна средняя температура гидросферы: -3,3 °С.
Главная закономерность влияния температуры на живые организмы связана с действием ее на скорость химических реакций. Величину температурного ускорения химических реакций удобно выражать коэффициентом Q10, показывающим, во сколько раз увеличится скорость реакции при повышении температуры на 10 °С:
Q 10 = (K1/ K2)/ ( t 2 -t 1 ) /10 ,
где K1, K2 — скорости реакции при температурах t1 и t2.
Для реакций абиотического характера Q10, в соответствии с правилом Вант-Гоффа, равен 2-3. Но в живых организмах химические реакции всегда идут с участием сложных ферментных систем, активность которых, в свою очередь, зависит от температуры. Поэтому Q10 в них на порядок отличается по амплитуде колебаний от такового в неживой природе, причем в обе стороны.
Важной экологической характеристикой являются температурные пороги жизни, т. е. интервалы температур, в пределах которых протекают жизненные процессы.
Максимальный температурный диапазон активной жизни (температура функционирующих клеток) несколько меньше диапазона жидкого состояния воды, а для большинства многоклеточных организмов – от 0 до +50 °С. Микроорганизмы эбулиофилы, обитающие в горячих поверхностных и глубинных источниках, могут сохранять способность к размножению при температуре до +75 °С, а некоторые архебактерии нормально существуют при +85…+105 °С. Диапазон температур, в котором клетки и многие организмы способны длительное время находиться в неактивном состоянии, существенно больше. Выживание при температуре намного ниже точки замерзания жидкостей тела (криобиоз) хорошо известно для семян и спор, отмечено у многих свободных клеток, низших беспозвоночных, некоторых рыб и амфибий, а также на изолированных тканях и органах высших животных и человека.
По обеспеченности наземных организмов теплом различают четыре основных термических пояса.
1. Тропический пояс. Средняя температура самого холодного месяца +15…+20 °С.
Амплитуда колебания температур не превышает 5 °С.
2. Субтропические пояса. Температура самого холодного месяца выше +4 °С, самого теплого — выше +20 °С. Возможны кратковременные морозы.
3. Умеренные пояса. Средняя длительность безморозного периода 70-80 суток.
Заморозки весной и осенью.
4. Холодные пояса. Заморозки возможны в течение всего вегетационного периода, продолжительность которого 1,5–2 месяца. Зима длинная, очень морозная. В каждом термическом поясе встречаются только такие организмы, пределы выносливости которых не выходят за рамки крайних значений температур, характерных для этих зон. Так, умеренные пояса заселены в основном эвритермными видами, тропический пояс – стенотермными теплолюбивыми видами, а холодные пояса - стенотермными холодолюбивыми видами.
По принципиальным особенностям теплового обмена различают две крупные экологические группы организмов: пойкилотермные и гомойотермные.
К пойкилотермным (от греч. роikilos – изменчивый, меняющийся, thermē – теплота, жар) организмам относят все таксоны органического мира, кроме двух классов позвоночных животных: птиц и млекопитающих. Принципиальная особенность теплообмена пойкилотермных организмов заключается в том, что благодаря относительно низкому уровню метаболизма главным источником поступления тепловой энергии у них является внешнее тепло. Именно этим объясняется прямая зависимость температуры тела пойкилотермных от температуры среды, точнее от притока теплоты извне, поскольку наземные пойкилотермные формы используют также и радиационный обогрев.
Впрочем, полное соответствие температур тела и среды наблюдается редко и свойственно главным образом организмам очень мелких размеров. В большинстве случаев существует некоторое расхождение между этими показателями. В диапазоне низких и умеренных температур среды температура тела организмов, не находящихся в состоянии оцепенения, оказывается более высокой, а в очень жарких условиях – более низкой. Причина превышения температуры тела над таковой среды заключается в том, что даже при низком уровне обмена продуцируется эндогенное тепло, которое и вызывает повышение температуры тела. Это проявляется, в частности, в существенном повышении температуры у активно двигающихся животных.
Например, у насекомых в покое превышение температуры тела над средой выражается десятыми долями градуса, тогда как у активно летающих бабочек, шмелей и других видов температура поддерживается на уровне +36…+40 °С даже при температуре воздуха ниже +10 °С.
Существует исключительный пример настоящей эффективной теплорегуляции у растений. Symplocarpus foetidus из семейства ароидных, цветущее ранней весной, для предотвращения замерзания соцветия-початка во время ночных заморозков включает ферментативный гидролиз крахмала в луковице, доставку глюкозы в початок и ее интенсивное ферментативное окисление, позволяющее поддерживать температуру соцветия на 10-15 °С выше температуры среды.
Пониженная по сравнению со средой температура при жаре свойственна наземным организмам и объясняется в первую очередь потерями тепла с испарением, которое при высокой температуре и низкой влажности существенно увеличивается.
Скорость изменений температуры тела пойкилотермов связана обратной зависимостью с их размерами. Это имеет большое экологическое значение, определяя для разных видов возможность заселения географических районов или биотопов с определенными режимами температур.
Но только настоящие «теплокровные», гомойотермные (от греч. homoios – одинаковый) животные – птицы и млекопитающие – могут поддерживать высокую температуру тела при значительных изменениях температуры среды. Они располагают совершенными нервными и гуморальными механизмами активной теплорегуляции, которые включают не только средства эффективной регуляции теплоотдачи, но и изменения интенсивности окислительных процессов и теплопродукции внутри организма.
Температурные адаптации пойкилотермных организмов. Известно немало видов растений, способных переносить низкие температуры, предварительно подготовившись к ним. Например, растения тропических лесов повреждаются при температуре +5…+8 °С, а обитатели сибирской тайги выдерживают полное промерзание. Такие организмы впадают в состояние глубокого покоя, или анабиоза, при котором все процессы жизнедеятельности сведены к минимуму. У растений в период подготовки к переходу в зимнее состояние идет накопление в клетках и тканях сахаров (до 20-30 %), а также некоторых аминокислот и других веществ, связывающих воду. Увеличивается вязкость протоплазмы и уменьшается содержание в ней свободной воды. Все это ведет к снижению точки замерзания жидкостей. У морозостойких деревьев в процессе сезонного холодового закаливания мембранные системы и белки в клетках видоизменяются таким образом, что межклеточная вода и ее замерзание не влияют на жизнестойкость клеток. Канадскими учеными показано, например, что озимые пшеницы в процессе холодового закаливания значительно изменяют состав растворимых белков. При этом образуется до восьми новых белков. Количество синтезируемых при закаливании белков коррелирует с морозоустойчивостью сорта. Такие процессы идут довольно медленно, поэтому для растений опасны быстрые похолодания и потепления, а особенно их чередования, так как клетки не успевают осуществить приспособительные перестройки.
Примерами морфологических адаптаций, касающихся формы растений холодных местообитаний, являются их небольшие размеры – карликовость (карликовые березы и ивы), стелющиеся формы (кедровый стланник, можжевельник, рябина), подушковидные формы растений (колючелистник, эспарцет, азорелла, сальсола).
Подушковидные растения привлекли внимание ботаников более 100 лет назад за их способность противостоять крайне неблагоприятным факторам внешней среды, не утратив элементов активности. Важным признаком подушковидных растений являются ничтожные миллиметровые приросты побегов. Растение существует десятки и сотни лет, оставаясь карликом.
Известны и другие морфологические приспособления растений к низким температурам, так называемые жизненные формы, классификацию которых, основанную всего на одном признаке – положении почек возобновления относительно земной поверхности, – предложил в 1903 г. датский ботаник К. Раункиер. Среди наземных растений он выделил пять главных типов, назвав их жизненными формами (рис. 4):
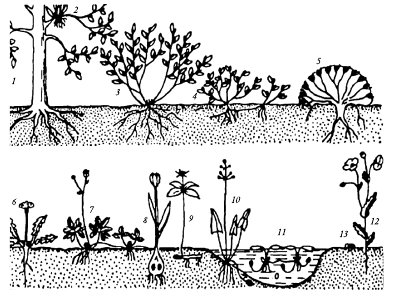
Рис. 4. Жизненные формы растений:
1—3 — фанерофиты; 4, 5 — хамефиты; 6, 7 — гемикриптофиты;
8—11 — криптофиты; 12 — терофит; 13 — семя с зародышем
фанерофиты (от греч. phaneros – видимый, phyton – растение) –растения, главным образом деревья и кустарники, у которых точки возобновления находятся на некотором расстоянии от поверхности почвы (выше 25 см);
хамефиты (от греч. chamoi – на земле) – растения (мелкие кустарники, кустарнички, низкорослые суккуленты), у которых почки возобновления расположены низко над поверхностью почвы (ниже 25 см);
гемикриптофиты (от греч. hemi- – полу-, kryptos – скрытый) – многолетние травянистые растения, у которых почки возобновления располагаются на уровне почвы и защищены отмершими листьями или снегом;
криптофиты – многолетние травы, у которых почки возобновления закладываются в луковицах, клубнях, корневищах и находятся в почве или под водой, благодаря чему они защищены от прямого воздействия среды;
терофиты (от греч. theros – лето) – однолетние травянистые растения, которые переживают неблагоприятный для их жизнедеятельности период в виде семян.
Выделенные категории хорошо отражают связь жизненных форм и экологических условий их существования, что подтверждают данные американского эколога Р. Уиттекера (1980). Фанерофиты абсолютно доминируют в тропических дождевых лесах (96 %), а также хорошо представлены в субтропических лесных формациях (65 %).
В тундре и степях начинают преобладать гемикриптофиты (60–63 %), а в пустынях господствуют терофиты (73 %) (рис. 5).
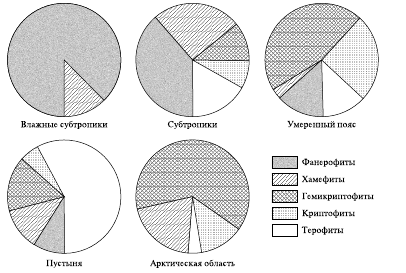
Рис. 5. Соотношение участия жизненных форм растений в местностях, расположенных в разных областях земного шара (по системе Раункиера)
Однако типы жизненных форм К. Раункиера слишком обширны и неоднородны, причем некоторые из них (хамефиты, гемикриптофиты) значительно представлены в различных ботанико-географических зонах (тундра, степи).
В зоне высоких температур при пониженной влажности (тропические и субтропические пустыни) сформировался своеобразный морфологический тип растений с незначительной листовой поверхностью или полным отсутствием листьев. У многих пустынных растений образуется беловатое опушение, способствующее отражению солнечных лучей и предохраняющее их от перегрева (акация песчаная, лох узколистный).
Интенсивность фотосинтеза при повышении температуры на 10 °С увеличивается в 2 раза, но лишь до +30…+35 °С, затем его интенсивность падает, и при +40…+45 °С фотосинтез вообще прекращается. При 50 °С большинство наземных растений погибают, что связано с интенсификацией дыхания растений при повышении температуры, а затем его прекращением при 50 °С.
Температура влияет и на ход корневого питания у растений: этот процесс возможен лишь при условии, когда температура почвы на всасывающих участках на несколько градусов ниже температуры наземной части растения. Нарушение этого равновесия влечет за собой угнетение жизнедеятельности растения и даже его гибель.
К физиологическим приспособлениям растений, сглаживающим вредное влияние высоких и низких температур, могут быть отнесены: интенсивность испарения – транспирации (от лат. trans – через, spiro – дышать, вдыхать); накопление в клетках солей, изменяющих температуру свертывания плазмы; свойство хлорофилла препятствовать проникновению солнечных лучей.
Активность эктотермных животных напрямую зависит от температуры. Низкие температуры для многих из них являются ограничивающим фактором, и поэтому способ выживания для большинства животных – резкое снижение метаболической активности (оцепенение, спячка). Местообитания с постоянно низкими температурами освоены лишь теми эктотермными организмами, которые в ходе длительной эволюции приобрели ряд адаптаций, препятствующих замерзанию внутри- и межклеточной воды, денатурации белков и изменению физического состояния липидов.
Переохлажденное состояние холодноводных рыб поддерживается накоплением в жидкостях тела так называемых биологических антифризов – гликопротеинов, понижающих точку замерзания и препятствующих образованию кристаллов льда в клетках и тканях. Концентрация этих веществ коррелирует с температурными условиями жизни. Так, у арктической трески она почти в 20 раз выше, чем у того же вида из умеренных широт.
У насекомых важную роль в переживании низких температур играет глицерин, также выступающий в качестве антифриза. У зимующих стадий ряда видов накопление в гемолимфе и тканях этого вещества снижает точку переохлаждения до -26…-37 °С и даже ниже. При этом во внеклеточной жидкости и гемолимфе образуются мелкие кристаллики льда или аморфные твердые структуры. Внутри клеток кристаллизация начинается лишь при крайне низких температурах (порядка -60 °С). Кроме того, глицерин, проникая в клетки, нормализует осмотическое давление. Накопление глицерина имеет хорошо выраженный сезонный характер: он отсутствует в тканях летом и в значительных количествах накапливается к зиме: у муравьев, например, до 10 %, у личинок Polygraphus – до 21 %, а у осы Braconcephi – даже до 30 %.
Помимо глицерина у насекомых и некоторых других беспозвоночных функционируют такие биологические антифризы, как низкомолекулярные (сахара) и высокомолекулярные (белки, гликоген), благодаря которым при акклиматизации к низким температурам повышается процент связанной воды. В тканях оцепеневших во льду молодых расписных черепах Chrysemis picta marginata обнаружены высокие концентрации глюкозы, глицерола и таурина, выполнявших функции криопротекторов.
Способность растений, животных и микроорганизмов переносить как низкие, так и высокие температуры повышается при обезвоживании тканей (сухая плазма выдерживает колебания температуры от -273 до +170 °С), и этот путь адаптации широко используется представителями разных таксонов. В опытах обезвоженные коловратки переносили замораживание при температуре -190 °С, высыхание – обычный для этой группы животных способ перенесения неблагоприятных по температуре и влажности сезонов. У многих пойкилотермных животных содержание воды в теле меняется сезонно, повышая холодостойкость в зимнее время. Особенно ярко это выражено у видов, подверженных зимой прямому действию низких температур, например у жуков-ксилофагов, зимующих в стволах деревьев. У видов, обитающих в менее жестких условиях, не столь отчетливо выражены и сезонные колебания содержания воды в тканях и холодостойкости соответственно.
Температура и развитие. Правило сумм температур. В наиболее генерализованной форме влияние температуры на обменные процессы прослеживается при изучении онтогенетического развития пойкилотермных организмов. Оно протекает тем быстрее, чем выше температура окружающей среды. Так, в экспериментах установлено, что развитие икры форели начинается при 0 °С. При температуре равной +2 °С личинки появляются через 205 дней, при +5 °С – через 82 дня, при +10 °С – через 41 день.
Во всех трех экспериментах произведение положительных температур среды на число дней развития остается постоянным: 410. Для осуществления генетической программы развития эктотермным организмам необходимо получить извне определенное количество тепла. Это тепло измеряется суммой эффективных температур. Под эффективной температурой понимается разница между температурой среды и температурным порогом развития. Порог развития и сумма эффективных температур для каждого вида определенные. Они зависят от приспособленности вида к условиям жизни.
Сумму эффективных температур рассчитывают по формуле
S = Д ⋅ (Т - t0),
где S – сумма тепла, или сумма эффективных температур, измеряемая в градусо-днях, градусо-часах, величина постоянная; Д – продолжительность развития; Т – температура окружающей среды; t0 – температурный порог развития (температура биологического нуля).
Например, у кузнечика Austroicetes cruciata при 20°С (т. е. на 4 °С выше порога) развитие яйца до момента вылупления занимает 17,5 сут, а при 30 °С (т. е. на 14 °С выше порога) – только 5 сут. Следовательно, при обеих температурах для завершения развития требуется 70 градусо-дней (или, точнее, «градусо-дней сверх порога развития»):
17,5 ⋅ 4 = 70 и 5 ⋅ 14 = 70.
Столько же требуется для развития кузнечика и при других температурах. Таким образом, произведение длительности развития на эффективную температуру есть величина постоянная.
Об эктотермах, в отличие от эндотермов, нельзя сказать, что для развития им требуется определенный промежуток времени. Им необходима некая комбинация времени и температуры, часто называемая физиологическим временем. Иначе говоря, время для развития эктотермов зависит от температуры, и если температура упадет ниже порога развития, то оно может «остановиться».
На организмы, обитающие в умеренных широтах, стимулирующее действие оказывает изменение температуры, крайне важное с экологической точки зрения. В. Шелфорд (1929) и Т. Парк (1948) в опытах, ставших хрестоматийными, показали, что в условиях переменной температуры, по сравнению с развитием при постоянной температуре, личинки и куколки яблоневой плодожорки формируются быстрее на 7–8 %, яйца – на 38 % и нимфы кузнечика – на 12 % . Позднее такая же закономерность была установлена для водных беспозвоночных.
Адаптации к температуре у эндотермных животных. Птицы и млекопитающие приобрели способность поддерживать температуру своего тела на постоянном уровне вне зависимости от температуры внешней среды за счет тепла, образуемого организмом в ходе метаболических процессов. Эндотермные организмы выработали ряд разнообразных приспособлений для терморегуляции. Так, благодаря толще подкожного жира и особенностям периферического кровообращения превосходно приспособлены к длительному пребыванию в ледяной воде многие ластоногие и киты. Песец, полярная сова и белый гусь легко переносят сильный холод без падения температуры тела и при поддержании разности температур тела и среды в 100 °С благодаря хорошо развитому меховому или перьевому покрову. При необходимости увеличение теплопродукции достигается повышением потребления пищи, окислением бурого жира, который служит исключительно источником тепла.
Увеличение теплоотдачи происходит путем потоотделения, уменьшения густоты шерстного либо перьевого покрова, расширения капилляров кожи, тепловой одышки или полипноэ.
В мире животных наблюдаются определенные морфологические адаптации, направленные на защиту организмов от неблагоприятного действия температур в различных климатических условиях.
Свидетельством этого может служить известное правило Бергмана (1847), согласно которому в пределах вида или достаточно однородной группы близких видов теплокровные организмы с более крупными размерами тела распространены в более холодных областях. Суть этого правила в необходимости поддержания для эндотермных животных постоянной внутренней температуры тела. Потери тепла происходят главным образом через поверхность тела: объем и вес его увеличиваются пропорционально кубу линейных размеров, а поверхность – пропорционально квадрату. Чем крупнее животное и, следовательно, меньше отношение поверхность – объем, тем меньше и потери тепла через теплоотдачу. Правило Бергмана хорошо подтверждается на пингвинах, медведях, тиграх и в лабораторных экспериментах (рис. 6, б).
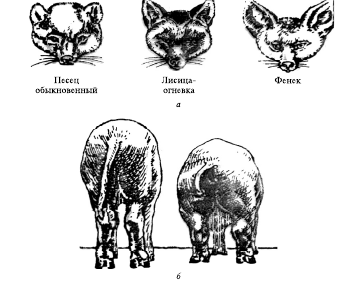
Рис. 6. Температурные адаптации у гомойтермных организмов:
а — генетически адаптированные к холоду лисицы (песцы) имеют маленькие уши, а адаптированные к теплу (фенек) обладают большими ушами. Различная величина ушей способствует более эффективной температурной регуляции. Теплоотдача уменьшается у песцов и увеличивается у лисиц из теплых климатических зон (правило Аллена); б — физиологическая (фенотипическая) адаптация к температурным условиям также способна изменить пропорции тела у развивающихся животных (правило Бергмана) (на рисунке изображены два животных одного помета (приплода): животное слева в течение семи недель содержалось при температуре 35 °С; животное справа — при температуре 5 °С; масса тела обоих животных совершенно одинакова)
Правило Аллена является следствием правила Бергмана. У млекопитающих, живущих в районах с холодным климатом, наблюдается значительное уменьшение поверхности ушей и хвоста, шея и лапы становятся короче, а формы тела – более приземистыми. Хорошим примером служат лисицы Старого Света: у фенека (Fennecus zerda) из жарких пустынь очень большие уши, у рыжей лисицы (Vulpes vulpes) наших широт они меньше, у песца (Alopex lagopus) – крошечные ушки и короткая морда (рис. 6, а).
Закон Глогера касается пигментации тела животных, обитающих в различных температурных условиях. Как правило, у видов из холодных и влажных зон отмечается более интенсивная, чем у обитателей сухих и теплых областей, пигментация тела (темная или темно-коричневая).
Существуют и радикальные формы защиты от холода – миграция в теплые края (перелеты птиц, спускание высокогорных обитателей на зиму на более низкие высоты и др.), зимовка –впадение в спячку на зимний период (сурок, белка, бурый медведь, летучие мыши). В последнем случае животные способны понижать температуру своего тела почти до нуля, замедляя метаболизм и тем самым трату запасенных питательных веществ. Но многие организмы умеренных широт в этот период ведут активный образ жизни (волки, олени, зайцы и др.), а некоторые даже размножаются (королевские пингвины и др.).
Темпертура тела около +37 °С выбрана в ходе эволюции высших животных не случайно, так как температурные зависимости многих химических реакций в этой точке температурной шкалы сближены и согласование их скоростей требует минимальных затрат энергии и молекулярной информации. Существенно, что температура вблизи +37 °С не только является оптимумом температуры тела высших животных, но и совпадает с предпочитаемой температурой многих наземных эктотермов.
В то же время почти все эндотермы плохо переносят значительное снижение температуры тела (гипотермию): ее падение ниже +10...+15 °С в большинстве случаев смертельно. Только зимоспячие млекопитающие (некоторые насекомоядные, рукокрылые и грызуны) при определенной физиологической готовности могут преодолевать барьер нижней летальной температуры и на длительное время впадать в состояние гипобиоза. Это позволяет им переживать время зимних холодов и бескормицы.
Состояние гипобиоза сближает зимоспячих с эктотермами, многие из которых приспособлены к переживанию неблагоприятного времени года в неактивном состоянии вплоть до анабиоза.
По сравнению с этими примерами приспособления растений и животных к высокой температуре более ограничены. Они допускают превышение температурного оптимума или нормальной температуры тела лишь на единицы градусов и основаны не столько на функциональных, сколько на морфологических и поведенческих механизмах.
Пойкилотермия и гомойотермия – выражение разных эволюционных стратегий теплообмена. Одна из них допускает широкое расселение и занятие многообразных экологических ниш на базе общей температурной толерантности. Это стратегия пойкилотермии.
Вторая стратегия – гомойотермия – обеспечивает не менее широкое расселение и существование в различных экологических условиях на основе поддержания теплоого гомеостаза внутренней среды. Благодаря ей обеспечивается сохранение высокой биологической активности практически во всем диапазоне переносимых температур, что связано с большими энергозатратами на процессы терморегуляции. Поэтому гомойотермия могла сформироваться только у высших животных, общий путь эволюции которых характеризовался направленным повышением метаболизма и ослаблением прямых зависимостей от внешних факторов путем повышения эффективности центральных интегрирующих и регуляторных систем организма. Было бы неправильно трактовать различия между этими группами как наличие каких-либо преимуществ у одной из них. Известно, что пойкилотермные организмы распространены по земному шару не менее широко, чем гомойотермные. Энергетическая стоимость температурных адаптаций у них ниже, чем у птиц и млекопитающих. С другой стороны, последние способны сохранять активность в широком диапазоне температур. Это дает им определенные преимущества в межвидовых отношениях, облегчая захват более выгодных экологических ниш. Поддержание постоянно высокого уровня метаболизма, вероятно, выгодно и на уровне экосистем, так как обеспечивает устойчивость биогенного круговорота. Не исключено, что в эволюции гомойотермии участие птиц и млекопитающих в поддержании стабильности функционирования биогеоценотических систем сыграло существенную роль.
7. Свет как экологический фактор
Как известно, свет – источник энергии для фотосинтеза и тепла для организмов. Однако этим его значение не исчерпывается, поэтому рассмотрим свет более подробно. В экологии под термином «свет» принято понимать весь диапазон излучения (в первую очередь солнечного) в пределах длин волн от 0,05 до 3000 нм (1 нанометр = 10-6 мм). Этот поток радиации распадается на несколько областей, отличающихся физическими свойствами и экологическим значением для живых организмов:
менее 150 нм –зона ионизирующей радиации;
150–400 нм – ультрафиолетовая (УФ) радиация;
400–800 нм – видимый свет (для разных организмов границы различаются);
800–1000 нм – инфракрасная (ИК) радиация;
1000–3000 нм – дальняя инфракрасная радиация.
Две последние области радиации несут в основном тепловую энергию, и их биологическое действие рассмотрено при обсуждении влияния температуры.
Не вся солнечная радиация достигает поверхности Земли. При прохождении через атмосферу часть ее рассеивается в воздухе и отражается, что влечет и качественные изменения исходного состава излучения. В частности, часть спектра с длинной волны до 300 нм отражается озоновым экраном.
Ионизирующая радиация. Она включает как космическое излучение, так и естественную и искусственную радиоактивность. В совокупности это главным образом естественный и техногенный радиоактивный фон. Ионизирующая радиация оказывает биологическое действие главным образом на субклеточном уровне (на ядро, митохондрии): в малых дозах – часто стимулирующее, в больших – повреждающее. «Порог» действия зависит от вида организма, ткани, клеток, а также длительности воздействия, его прерывистости и др. Известен и мутагенный эффект. В целом, влияние этого излучения на организмы не достаточно изучено и составляет предмет радиобиологии.
Ультрафиолетовые лучи (УФЛ). По длине волны подразделяются на УФ-С (150–280 нм), УФ-В (280–320 нм) и УФ-А (320–400 нм). УФ-С весьма опасены, так как интенсивно адсорбируются кожей. Однако они практически полностью поглощаются озоновым экраном. УФ-В обладают канцерогенным действием неизученного до конца механизма. Существует основание утверждать, что они связаны с нарушением структуры молекул ДНК. Кроме того, известно, что эти лучи инактивируют в коже клетки Лангерганса, отвечающие за ее иммунитет. УФ-В также большей частью поглощаются озоновым экраном.
До поверхности земли доходят большей частью лучи с длинной волны от 300 нм. Они обладают довольно большой энергией и оказывают на организмы главным образом химическое действие, в частности стимулируя процессы клеточного синтеза. Облучение ультрафиолетом 320–350 нм повышает продуктивность молодняка свиней и успешно используется в интенсивном животноводстве. Под действием УФЛ в организме синтезируется витамин D, регулирующий обмен Са и Р, а соответственно, рост и развитие скелета. Особенно велико значение этого витамина для растущего молодняка. Поэтому, например, лисицы и барсуки, выводящие детенышей в норах, солнечным утром выносят их на освещенные места у норы.
Слишком сильное облучение УФЛ вредно для организма, так как повреждает активно делящиеся клетки. Как приспособление к экранированию организма от передозировки УФ у животных формируются темные пигменты, поглощающие эти лучи. Такова природа загара у человека, пигментации верхнего полюса икринок у лягушек, верхних оболочек головного мозга и мошонки у пустынных сусликов (защищены нервные и половые клетки).
Видимый свет. Составляет 40–50 % солнечной энергии, достигающей Земли. Его важнейшее экологическое значение определяется тем, что свет является фактором фотосинтеза, выступая здесь в роли источника энергии. Но в фотосинтезе участвует лишь часть спектра с длиной волны в пределах от 380 до 760 нм, которую называют областью физиологически активной радиации (ФАР). Внутри нее для фотосинтеза наибольшее значение имеют красно-оранжевые (600–700 нм) и фиолетово-голубые (400–500 нм), наименьшее – желто-зеленые (500–600 нм) лучи. Последние отражаются, что и придает хлорофиллоносным растениям зеленую окраску. Способность воспринимать лучистую энергию у хлорофилла и зрительных пигментов животных очень близка, поэтому область фотосинтетически активной радиации практически совпадает с диапазоном видимой части спектра (400–700 нм). Лишь бактериохлорофилл способен поглощать свет с длиной волны до 900 нм. Зеленые растения поглощают ~75 % падающей на них лучистой энергии доступного спектра, но на фотосинтез используется только 1–10 % ее. Остальная энергия затрачивается на транспирацию и другие процессы. Разнообразие световых условий на нашей планете чрезвычайно велико: от таких сильно освещенных территорий, как высокогорья, пустыни, степи, до сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т.д. Соответственно разнообразны и механизмы приспособления растений к жизни при том или ином световом режиме.
Растения по отношению к освещенности подразделяются на светолюбивые (гелиофиты), теневые (сциофиты) и теневыносливые. Первые две группы обладают разными диапазонами толерантности в пределах экологического спектра освещенности. Яркий солнечный свет – оптимум гелиофитов (луговые травы, хлебные злаки, сорняки и др.), слабая освещенность – оптимум теневых (растения таежных ельников, лесостепных дубрав, тропических лесов). Первые не выносят тени, вторые – яркого солнечного света.
Теневыносливые растения имеют широкий диапазон толерантности к свету и могут развиваться как при яркой освещенности, так и в тени.
У животных световой фактор прежде всего играет огромную роль в зрительной ориентации в пространстве. Практически у всех групп животных, начиная с подвижных форм кишечнополосных – медуз, развиваются светочувствительные органы – глаза, способные различать спектральный состав света. Особенно велика их роль для животных, обитающих в наземно-воздушной среде. Именно у них орган зрения достигает наибольшего совершенства.
Ориентация животных на свет осуществляется в результате фототаксисов: положительного (движение в сторону большей освещенности) или отрицательного (движение в сторону наименьш
Дата добавления: 2022-04-12; просмотров: 404;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
