ЭКОЛОГИЧЕСКИХ ФАКТОРОВ НА ОРГАНИЗМ
В характере воздействия факторов среды и ответных реакциях живых организмов выявлен ряд общих закономерностей, которые укладываются в некоторую общую схему действия экологического фактора разной дозы на жизнедеятельность организмов.
Количественное выражение экологического фактора в пределах зоны толерантности определяется в основном значениями, представленными тремя кардинальными точками – минимума, оптимума и максимума, и на рис. .2 кривая 1 имеет вид куполообразной кривой, так называемой кривой толерантности. Крайние пороговые значения (точки минимума и максимума) называются нижним и верхним пределами выносливости.
Зона, непосредственно прилегающая к точке оптимума, называется зоной оптимума или зоной комфорта. В этой зоне организм максимально адаптирован к действию экологического фактора, и количество последнего соответствует экологическим потребностям организма. Значение оптимума не является абсолютной величиной для конкретного вида, а зависит от стадии онтогенеза, периода жизни и действия других факторов. Зона, прилегающая к зоне оптимума, называется зоной нормы. Ей соответствует такое количество экологического фактора, при котором все жизненно важные процессы протекают нормально, однако для поддержания их на этом уровне необходимы дополнительные энергетические затраты.
В зоне пессимума нормальный ход процессов жизнедеятельности затруднен. Повторяемость описанных тенденций позволяет рассматривать их как фундаментальный биологический принцип: для каждого вида растений и животных существует оптимум, зона нормальной жизнедеятельности, стрессовые зоны и пределы выносливости в отношении каждого фактора среды.
Адаптация к каждому фактору связана с затратами энергии. В зоне оптимума адаптивные механизмы отключены и энергия расходуется только на фундаментальные жизненные процессы (энергозатраты на базальный метаболизм). При выходе значений фактора за пределы оптимума включаются адаптивные механизмы, функционирование которых сопряжено с определенными затратами энергии – тем большими, чем дальше значение фактора отклоняется от оптимального (рис.2, кривая 2). При этом усиление энергорасходов на адаптацию ограничивает возможный набор форм жизнедеятельности организма: чем дальше от оптимума находится количественное выражение фактора, тем больше энергии направленно расходуется на адаптацию и тем меньше «степеней свободы» в проявлении иных форм деятельности. В конечном итоге нарушение энергетического баланса организма наряду с повреждающим действием недостатка или избытка фактора ограничивает диапазон переносимых им изменений. Размах адаптивных изменений количественного выражения фактора определяется как экологическая валентностьили экологическая пластичность вида по данному фактору. Величина ее различна у разных видов.
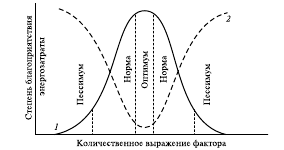
Рис. 2. Влияние изменения количественного выражения
фактора среды на жизнедеятельность организма:
1 — степень благоприятствования данных доз для организма;
2 — величина энергозатрат на адаптацию (схема условная; предполагается, что все остальные факторы действуют в оптимуме
Экологически непластичные, т. е. маловыносливые виды, для существования которых необходимы строго определенные, относительно постоянные условия внешней среды, называются стенобионтными (от греч. stenos – узкий, bios – жизнь), а те, которые могут жить в широком диапазоне изменчивости условий среды, – эврибионтными (от греч. eurys – широкий) (рис. 3).
В зависимости от конкретного фактора среды различают организмы стено- и эври-термные по отношению к температуре, стено- и эврифотные по отношению к свету, стено- и эврибатные по отношению к давлению, стено- и эвригалинные по отношению к концентрации солей. Важно подчеркнуть, что явление стенобионтности фактически используется в практике экологической индикации качества окружающей среды. Узкоспециальные по отношению к ряду факторов видовые популяции могут служить более чувствительными показателями качества среды, чем физические и химические.
Экологическая валентность как видовое свойство эволюционно формируется в качестве приспособления к той степени колебаний данного фактора, которая свойственна естественным местам обитания вида. Поэтому, как правило, переносимый данным видом диапазон колебаний фактора соответствует его естественной динамике: обитатели континентального климата выдерживают более широкие колебания температуры, чем жители приэкваториальных муссонных регионов. Сходные отличия обнаруживаются и на уровне различных популяций одного вида, если они занимают не одинаковые по условиям места обитания.
Помимо величины экологической валентности, виды (и популяции одного вида) могут отличаться и местоположением оптимума на шкале количественных изменений фактора. Виды, приспособленные к высоким дозам данного фактора, терминологически обозначаются окончанием -фил (от греч. phyleo – любить): термофилы (теплолюбивые виды), оксифилы (требовательны к высокому содержанию кислорода), гигрофилы (населяют места с высокой влажностью) и т. д. Виды, обитающие в противоположных условиях, обозначаются термином с окончанием -фоб (от греч. phobos – страх): галлофобы – обитатели пресных водоемов, не переносящие осолонения, хианофобы – виды, избегающие глубокоснежья, и т. п. Нередко такие формы характеризуют «от обратного»: например, виды, не переносящие избыточного увлажнения, чаще называют ксерофильными (сухолюбивыми), чем гигрофобными; подобным же образом взамен термина «термофоб» чаще употребляют «криофил» (холодолюбивый).
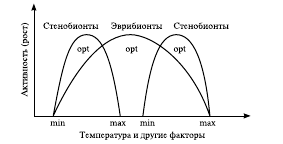
Рис. 3. Экологическая пластичность видов
Информация об оптимальных значениях отдельных факторов и диапазоне переносимых ими колебаний достаточно полно характеризует отношение вида (популяции) к каждому исследованному фактору. Следует, однако, иметь в виду, что рассмотренные категории дают лишь общее представление о реакции вида на воздействие отдельных факторов. Это важно при общей экологической характеристике вида и решении ряда прикладных задач экологии (например, проблема акклиматизации вида в новых условиях), хотя и не определяет полного объема взаимодействия вида с условиями среды в сложной природной обстановке.
В совокупности условий существования всегда можно выделить фактор, который сильнее других влияет на состояние организма или популяцию. Так, дефицит одного из важных ресурсов (вода, свет, пища, незаменимая аминокислота) будет ограничивать жизнедеятельность даже тогда, когда все остальные условия оптимальны. Фактор, который при определенном наборе условий окружающей среды ограничивает какое либо проявление жизнедеятельности, называется лимитирующим. Понятие лимитирующего фактора связано с законом минимума Либиха. Еще в середине XIX в. известный немецкий химик Ю. Либих, разрабатывая систему применения минеральных удобрений, сформулировал правило минимума, в соответствии с которым возможность существования данного вида в определенном районе и степень его «процветания» зависят от факторов, представленных в наименьшем количестве. Ученый установил, что урожай зерна часто лимитируется не теми питательными веществами, которые требуются в больших количествах (СО2, Н2О и др.), поскольку они, как правило, присутствуют в изобилии, а теми, которые необходимы в малых количествах и которых в почве недостаточно. Классическими примерами воздействия лимитирующего фактора на развитие растений являются исчерпание запасов бора в почве в результате возделывания одной и той же культуры в течение длительного времени или количество доступной влаги в засушливых аридных районах.
Позднее действие закона минимума Либиха было дополнено двумя принципами. Первый – ограничительный: закон может быть применим лишь в условиях стационарного состояния, т. е. когда приток и отток энергии и веществ сбалансированы.
Второй принцип – взаимодействие различных факторов. Например, некоторым растениям нужно меньше цинка, если они растут не на ярком солнечном свету, а в тени; значит, концентрация цинка в почве с меньшей вероятностью может быть лимитирующей для растений в тени, чем для растений на свету.
Лимитирующим может быть не только недостаток (минимум), но и избыток (максимум) экологического фактора. Представление о лимитирующем влиянии максимума наряду с минимумом развил американский зоолог В. Шелфорд в 1913 г.
Закон толерантности Шелфорда: лимитирующим фактором процветания может быть как минимум, так и максимум экологического фактора, диапазон между которыми определяет величину толерантности, выносливости организма к данному фактору.
Интересный пример действия закона толерантности В. Шелфорда (много «хорошо» — тоже нехорошо) приводит Ю. Одум (1986). Создание утиных ферм вдоль рек, впадающих в южную бухту в проливе Лонг-Айленд близ Нью-Йорка, стало причиной сильного удобрения вод утиным пометом, из-за чего значительно увеличилась численность фитопланктона и, самое главное, произошла его структурная перестройка: дино флагелляты и диатомовые водоросли Nitzschia оказались почти полностью заменены зелеными жгутиковыми, относящимися к родам Nannochloris и Stichococcus.
Знаменитые голубые устрицы, ранее процветавшие на рационе из традиционного фитопланктона и бывшие предметом выгодного водного хозяйства, постепенно исчезли, не адаптировавшись к новому виду пищи. Таким образом, избыток биогенов оказал лимитирующее воздействие на устриц.
Существует ряд вспомогательных принципов, дополняющих «закон толерантности».
1. Организмы могут иметь широкий диапазон толерантности в отношении одного фактора и узкий диапазон в отношении других факторов.
2. Организмы с широким диапазоном толерантности ко всем факторам обычно наиболее широко распространены. Например, карась, карп и многие другие рыбы переносят довольно низкое (менее 2 мг/л) содержание кислорода в воде, большую ее мутность, высокий диапазон температур. Поэтому они широко распространены в водоемах разных типов. Форель, напротив, встречается в реках, где концентрация кислорода более 2 мг/л. При содержании кислорода менее 1,6 мг/л она гибнет.
3. Если условия по одному экологическому фактору не оптимальны для вида, то может сузиться и диапазон толерантности к другим экологическим факторам. Например, при недостатке азота снижается засухоустойчивость злаков, т. е. растениям для выживания требуется больше воды.
4. В природе организмы часто оказываются в условиях, не соответствующих оптимальному диапазону того или иного фактора, определенному в лаборатории. В этом случае для жизнедеятельности организма более важным оказывается иной фактор. Например, некоторые тропические орхидеи в лаборатории при невысоких температурах лучше развиваются на солнце, чем в тени. В природе же они растут исключительно в тени, так как не переносят воздействия прямых солнечных лучей.
5. Периоды размножения обычно являются критическими для организмов. Многие факторы среды в это время становятся лимитирующими. Пределы толерантности для размножающихся особей и зародышей обычно у́же, чем для неразмножающихся взрослых животных и растений. Взрослые голубые крабы рода Portunus хорошо переносят солоноватую и пресную воду с высоким содержанием хлоридов, поэтому часто заходят в реки вверх по течению, однако не размножаются, так как для личинок крабов нужна высокая соленость. Зрелый кипарис способен расти как на сухом нагорье, так и на полностью заливаемой почве, тогда как для прорастания семени требуется увлажненная, но не заливаемая почва. Географическое распространение промысловых птиц часто определяется влиянием климатических факторов на стадиях раннего онтогенеза, а не на взрослых особей. К недостатку пищи более устойчивы взрослые особи. Таким образом, в течение индивидуального развития (онтогенеза) реакция животных и растений на экологические факторы меняется.
Большинство экологических факторов – температура, влажность, ветер, наличие пищи, хищники, паразиты, конкуренты и т. д. –отличаются значительной изменчивостью во времени и пространстве.
Изменения факторов среды наблюдаются в течение года и суток, в случае приливов и отливов в океане, при бурях, ливнях, обвалах, похолодании или потеплении климата.
Степень изменчивости каждого из этих факторов зависит от особенностей среды обитания. Например, температура сильно варьирует на поверхности суши, но почти постоянна на дне океана или в глубине пещер. Паразиты млекопитающих живут в условиях избытка пищи, тогда как для большинства хищников ее запасы все время меняются вслед за изменением численности жертв.
Один и тот же фактор среды имеет разное значение в жизни совместно обитающих организмов. Например, солевой состав почвы важен для минерального питания растений, но безразличен для большинства наземных животных.
Взаимодействие факторов в комплексах. Совокупное действие на организм нескольких факторов среды обозначают термином «констелляция». Экологически важно то обстоятельство, что констелляция не представляет собой простой суммы влияния факторов: при комплексном воздействии между отдельными факторами устанавливаются особые взаимодействия, когда влияние одного фактора в какой-то мере изменяет (усиливает, ослабляет и т. п.) характер воздействия другого.
Известно, например, что реакции газообмена у рыб существенно отличаются в условиях разной солености воды. В некоторых случаях недостаток одного фактора частично компенсируется усилением другого. Явление частичной взаимозаменяемости действия экологических факторов называется эффектом компенсации. Ю. Одум (1975) приводит такой пример: некоторые моллюски (в частности, Mytilus galloprovincialis) при отсутствии или дефиците кальция могут строить свои раковины, частично заменяя кальций стронцием при достаточном содержании в среде последнего. В пустынях недостаток осадков в определенной мере восполняется повышенной относительной влажностью воздуха в ночное время. Так, в пустыне туманов Намиб (Африка) среднегодовое количество осадков составляет примерно 30 мм, а с росой за 200 дней с туманом дополнительно поступает 40–50 мм осадков в год.
Климатические факторы могут замещаться биотическими (вечнозеленые виды южных растений в более континентальном климате могут расти в подлеске под защитой верхних ярусов, создавая собственный биоклимат). Такая компенсация факторов обычно создает условия для физиологической акклиматизации вида – эврибионта, имеющего широкое распространение. Акклиматизируясь в данном конкретном месте, он создает своеобразную популяцию, экотип, пределы толерантности которой соответствуют местным условиям.
Однако полное отсутствие в среде фундаментальных экологических факторов (физиологически необходимых: света, воды, углекислого газа, питательных веществ) не может быть компенсировано (заменено) другими факторами.
Экологические факторы действуют на живые организмы по-разному. Они могут выступать как раздражители, вызывающие приспособительные изменения физиологических функций; ограничители, обусловливающие невозможность существования в данных условиях; модификаторы, вызывающие морфологические и анатомические изменения организмов. Таким образом, воздействие экологических факторов на конкретные организмы способно:
1) устранять те или иные виды с той или иной территории
2) изменять конкурентноспособность видов и приводить к перестройкам в сообществах разных типов;
3) вызывать появление адаптивных изменений у видов;
4) через воздействие на отдельные виды оказывать существенное влияние на биогеохимические циклы в биосфере.
СРЕДЫ ЖИЗНИ
Первой средой жизни, освоенной организмами, была водная среда, или гидросфера.
Это самая обширная область, занимающая до 71 % площади нашей планеты. Основное количество воды (97 %) сосредоточено в морях и океанах и лишь менее 0,5 % находится в реках, озерах, болотах. Большая же часть пресной воды заключена в ледниках.
В водной среде обитает около 150 тыс. видов животных и более 10 тыс. видов растений, называемых гидробионтами. Главным фактором, определяющим условия передвижения гидробионтов и создающим давление на разных глубинах, является плотность воды. Для дистиллированной воды она равна 1 г/см3 при +4 °С, а при содержании растворенных солей может достигать 1,35 г/см3. На плотность пресной воды сильное влияние оказывает температура: она наибольшая при температуре +4 °С. При повышении или понижении температуры плотность воды уменьшается. При замерзании вода расширяется, увеличивая свой объем, и становится легче. Благодаря этому свойству лед располагается на поверхности водоема, в то время как наиболее плотная жидкая вода с положительной температурой находится подо льдом.
При активном передвижении гидробионты преодолевают сопротивление плотной воды за счет обтекаемой торпедообразной формы их тела. Вместе с тем высокая плотность воды и ее выталкивающая сила создают возможность опоры на нее. Поэтому в толще водной среды выделяют особые экологические группировки гидробионтов:
планктон (пассивно «парящие» организмы) и нектон (активно плавающие и способные преодолевать течения). Большинство рыб, некоторые беспозвоночные животные и цианобактерии имеют гидростатические аппараты (плавательный пузырь, газовые вакуоли и др.), позволяющие им флотировать в толще воды и «зависать» в ней на определенных глубинах. Благодаря способности воды удерживать в ее толще живые организмы (фито-, зоо-, бактериопланктон) и мертвую органическую взвесь у многих водных животных (подвижных, малоподвижных и прикрепленных) развился особый способ добычи пищи – фильтрационный. Высокая плотность воды создает возрастающее с глубиной давление, равное примерно 1 атм. на каждые 10 м. Температурный режим водоемов более устойчив, чем на суше. Это связано с физическими свойствами воды и, прежде всего, с высокой величиной удельной теплоемкости. Чтобы изменить температуру 1 г воды на 1 °С, нужно затратить 4,19 Дж тепла (в 500 раз больше, чем для воздуха). Благодаря этому свойству вода, медленно нагреваясь и остывая, уменьшает амплитуду суточных и сезонных колебаний температур, стабилизируя ее. Так, амплитуда годовых колебаний температуры в верхних слоях океана не более 10-15 °С, а в континентальных водоемах — 30–35 °С. Глубокие слои водоема имеют постоянные и более низкие величины температур. В экваториальных водах среднегодовая температура поверхностных слоев +26-27 °С, в полярных — около 0 °С и ниже. Более устойчивый температурный режим водоемов по сравнению с наземно-воздушной средой сформировал стенотермность большинства населяющих их гидробионтов. Эвритермные виды встречаются главным образом в мелких континентальных водоемах и литоральной зоне морей высоких и умеренных широт, где значительны суточные и сезонные колебания температуры.
Вода обладает большой скрытой теплотой плавления: для превращения 1 г льда в воду без изменения температуры необходимо затратить 80 кал. У воды самая высокая из известных скрытая теплота парообразования. При испарении 1 г воды поглощается 537 кал. Благодаря этому свойству происходит смягчение климата.
Вода – хороший растворитель разнообразных минеральных веществ. В зависимости от концентрации растворенных в ней солей выделяют пресные (до 0,5 г/л), солоноватые (0,5—16 г/л), морские (16—47 г/л) и пересоленные (47–350 г/л) воды. Заселение организмами водоемов с разной соленостью связано напрямую с их способностью к осморегуляции. Большинство гидробионтов являются стеногалинными организмами.
С повышением солености возрастает плотность воды и понижается температура ее замерзания. В воде растворяются и газы. Однако кислорода в ней содержится в 30 раз меньше, чем при той же температуре в равном объеме воздуха, тогда как углекислого газа, в воде больше, чем в воздухе. Содержание кислорода и углекислого газа в водоемах сильно изменится в течение суток: в светлое время содержание кислорода в воде повышается, а углекислого газа понижается вследствие осуществления фотосинтеза фотоавтотрофными гидробионтами; в ночное время происходит противоположное явление. Коэффициент диффузии кислорода в воде примерно в 320 тыс. раз ниже, чем в воздухе. В водоемах обогащение кислородом происходит за счет фотосинтетической аэрации и диффузии из воздуха. Диффузии способствуют ветер и движение воды. При повышении температуры воды, снижающей растворимость кислорода, отсутствии циркуляции воды за счет ветрового перемешивания в слоях, сильно заселенных живыми организмами, а также богатых мертвым органическим веществом в донной области водоемов, может создаваться резкий дефицит кислорода, особенно в ночное время, приводящий к гибели водных организмов — замору. Вследствие этого кислород в воде является лимитирующим фактором для жизни гидробионтов. Свет проникает в толщу водоемов на разную глубину в зависимости от содержания в ней минеральных и органических взвешенных и растворенных веществ, а также угла наклона солнечных лучей, падающих на поверхность воды. Поэтому прозрачность природных вод невелика и находится в пределах от 0,1 до 66,5 м (величину прозрачности определяют путем погружения закрепленного на тросе белого диска Секки в воду до предельной глубины его видимости). Самые прозрачные воды в Саргассовом море – 66,5 м, в мелких морях прозрачность составляет 5–15 м, в реках — 1–1,5 м. Нижняя граница прозрачности по диску Секки соответствует 5 % падающего на поверхность солнечного излучения. Фотосинтез продолжает идти и при меньшей освещенности, однако пятипроцентный уровень соответствует нижней границе основной фотосинтетической (эуфотической) зоны. Граница зоны фотосинтеза поэтому сильно варьирует в разных водоемах. В самых чистых водах эуфотическая зона простирается до глубин не ниже 200 м, сумеречная, или дисфотическая, зона занимает глубины до 1000–1500 м, а более глубокая афотическая зона полностью лишена солнечного света.
Количество света в верхних слоях водоемов сильно меняется и зависит от широты местности, а также времени года. Так, например, длинные полярные ночи, наличие ледяного покрова на водоемах сильно ограничивают время, пригодное для фотосинтеза.
Световые лучи с разной длиной волны поглощаются неодинаково: красные поглощаются уже в поверхностных слоях водоема, в то время как синие и в особенности зеленые части солнечного спектра проникают значительно глубже. Соответственно сменяют друг друга с глубиной зеленые, бурые и красные водоросли, имеющие разные специализированные пигменты для улавливания света с разной длиной волны.
Наземно-воздушная среда жизни освоена в ходе эволюции значительно позднее, чем водная. Она является наиболее разнообразной как во времени, так и в пространстве. Тела живых организмов окружены воздухом — газообразной подвижной средой с низкой плотностью (в 800 раз меньше, чем у воды), невысоким и постоянным давлением (около 760 мм рт. ст.), высоким содержанием кислорода и малым количеством водяных паров. Это сильно изменяет условия дыхания, водообмена и передвижения живых существ.
Низкая плотность воздуха определяет его малую подъемную силу и незначительную опорность. Поэтому наземные организмы имеют в своем теле хорошо развитые механические ткани и опору на поверхность земли. Низкое сопротивление воздуха при движении позволяет животным передвигаться со значительно большей скоростью, чем гидробионтам.
Жизнь во взвешенном состоянии в воздухе невозможна. Лишь микроскопические организмы, пыльца, семена, споры временно присутствуют в воздухе и переносятся воздушными потоками, с помощью которых они расселяются. Определенные животные (насекомые, птицы, рукокрылые) способны к активному полету. Однако они используют его только для расселения и поиска пищи. Все остальные функции осуществляются на поверхности земли.
Газовый состав воздуха в приземном слое атмосферы довольно однороден и стабилен (азот – 78 %, кислород – 21 %, аргон – 0,9 %, углекислый газ – 0,03 % по объему) благодаря высокой диффузионной способности газов и постоянному его перемешиванию конвекционными и ветровыми потоками. Высокое содержание кислорода способствовало повышению обмена веществ у наземных организмов по сравнению с первичноводными. Именно в наземной среде на базе высокой эффективности окислительных процессов в организме возникла гомойотермия животных (у птиц и млекопитающих). Кислород из-за постоянно высокого его содержания в воздухе не лимитирует жизнь в наземной среде.
Режимы влажности на суше очень разнообразны — от полного и постоянного насыщения воздуха водяными парами в некоторых районах тропиков до практически полного их отсутствия в сухом воздухе пустынь. Велика также суточная и сезонная изменчивость содержания водяных паров в атмосфере. Наземные организмы постоянно сталкиваются с проблемой потери воды. Эволюция наземных организмов проходила в направлении приспособления к добыванию и сохранению влаги.
Свет является источником энергии для фотосинтеза и тепла. Наземные растения используют в процессе фотосинтеза электромагнитные волны главным образом синей и красной части видимой области солнечного спектра (390–760 нм). Интенсивность и количество света в наземно-воздушной среде наиболее велики и практически не лимитируют жизнь зеленых растений. Для подавляющего большинства животных с дневной и даже ночной активностью зрение играет важную роль в ориентации, поиске добычи, способов маскировки и т. п.
Рельеф местности и свойства грунта оказывают существенное влияние на жизнь наземных организмов, формируя особенности светового, температурного режимов, влажности.
Большой размах температурных колебаний в сочетании с различным режимом влажности, облачности, осадков, силы и направления ветра создает большое разнообразие погодных условий, действию которых подвергаются организмы. В различных географических районах складываются сходные погодные условия, формирующие их климат.
Для большинства наземных организмов (в особенности мелких) каждой климатической зоны важны также условия их непосредственного местообитания, зависящего от особенностей рельефа, экспозиции, наличия растительности, что в совокупности формирует микроклимат. Например, температура поверхности дерева, обращенной к югу, будет намного выше, чем на северной. Резко отличаются температура, влажность, сила ветра, освещенность на открытых пространствах и в лесу, а зимой – на открытых участках почвы и под снегом, слоем опавших листьев, в норах, дуплах, пещерах и т. п.
Разнообразие микроклиматов создало гораздо больше вариантов условий в наземновоздушной среде, что способствовало возникновению в ходе эволюции более значительного количества видов наземных организмов по сравнению с водными.
Почва представляет собой сложную систему, состоящую из твердых минеральных частиц и органических остатков (перегноя), окруженных воздухом и водой. В зависимости от типа почвы – глинистая, песчаная, глинисто-песчаная и др. – она в большей или меньшей степени пронизана полостями, заполненными смесью газов и водными растворами. В почве, по сравнению с приземным слоем воздуха, сглажены температурные колебания, а на глубине 1 м не ощутимы и сезонные изменения температуры. Верхний горизонт почвы содержит определенное количество перегноя (гумуса), от которого зависит продуктивность растительного покрова. Расположенный под ним средний слой включает вымытые из верхнего слоя и преобразованные вещества. Нижний слой представлен материнской породой.
Вода в почве находится в крупных полостях, куда проникает под действием силы тяжести (гравитационная), либо в виде пленочной воды, удерживаемой поверхностью почвенных частиц. Вода содержится также в мельчайших пространствах – порах и может передвигаться по ним в разных направлениях, подобно движению ее в капилляре. В почвенном воздухе присутствует и парообразная вода.
Состав почвенного воздуха резко меняется с глубиной: содержание кислорода уменьшается, а углекислого газа – возрастает. При затоплении почвы водой или интенсивном гниении органических остатков возникают бескислородные зоны. По комплексу абиотических условий почву можно рассматривать как среду, имеющую многие общие признаки сходства как с водной, так и наземно-воздушной средами, а также собственные отличительные признаки.
Практически все живые организмы служат средой обитания для других организмов — паразитов. Паразитические организмы на протяжении всей своей жизни или части ее (паразитоиды) пользуются органическими веществами другого организма, называемого хозяином. Таким образом, хозяин является средой жизни паразита. Паразит может поселиться на поверхности тела хозяина (эктопаразитизм) или внутри него (эндопаразитизм).
В отличие от свободноживущих организмов, непосредственно сталкивающихся и взаимодействующих с факторами внешней среды, паразиты перекладывают бремя регуляции отношений с нею на организм хозяина. Также через хозяина регулируется получение паразитом пищи, являющейся важнейшим фактором внешней среды и выживания паразита.
Известный русский паразитолог академик Е. Н. Павловский различал двоякую среду паразитов: среду первого порядка – непосредственное место обитания, т. е. организм хозяина, и среду второго порядка – условия, окружающие хозяина. Не следует считать, что окружающая паразита среда постоянна. Она может резко меняться на протяжении короткого промежутка времени (неделя, день), например при заболевании хозяина, выработки им иммунитета и т. п. Для освоения этой среды – организма хозяина – у паразитов в ходе эволюции выработался ряд адаптаций: органы прикрепления к телу хозяина, защитные покровные образования, высокая плодовитость, сложные циклы развития со сменой хозяина и др.
Дата добавления: 2022-04-12; просмотров: 374;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
