При этом величина, форма, и длительность отдельного импульса не зависят от природы и силы раздражителя.
Аксоны могут возбуждаться электрическими импульсами различной формы, теплом, холодом, химическим воздействием и механическим напряжением – все это приводит к одинаковому эффекту. Скорость распространения нервного импульса была измерена уже Гельмгольцем. При сравнимых диаметрах аксонов она больше у миелинизированных волокон (у кошки: 0.7…2.3 м/с –немиелинизированные волокна, 10…100м/с - миелинизированные волокна).
В лабораториях потенциал действия обычно возбуждают с помощью электрического тока, так как при этом легко контролировать силу, время и длительность раздражителя. Типичная схема эксперимента показана на рис.2 (слева).
Процесс нервного возбуждения развивается следующим образом Если пропустить через аксон слабый импульс тока Io, приводящий к деполяризации мембраны, то после снятия внеш. воздействия Io (1a) потенциал  (1б) монотонно возвращается к исходному уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая из конденсатора и пост. сопротивления.
(1б) монотонно возвращается к исходному уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая из конденсатора и пост. сопротивления.
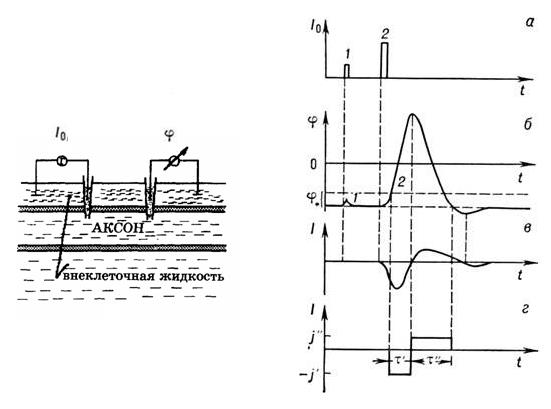
Рис. 2. Развитие потенциала действия в нервном волокне:а- подпороговое (1) и надпороговое (2)раздражения;б-мембранный отклик; при надпороговом раздражении проявляется полный потенциал действия 2б;в- ионный ток, протекающий черезмембрану при возбуждении;г- аппроксимацияионного тока в простой аналитической модели.
Если импульс тока превышает нек-рую пороговую величину, потенциал продолжает изменяться и после выключения возмущения; потенциал становится положительным и только потом возвращается к уровню покоя, причём вначале даже несколько проскакивает его (область гиперполяризации, рис. 2). Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом действия. Одновременно через мембрану течёт ионный ток, направленный сначала внутрь, а потом наружу (рис. 2,в).
Классические эксперименты такого рода были выполнены Ходжкиным и Хаксли (1952г).Они исследовали распространение нервного импульса в гигантских аксонах кальмаров (диаметр -600 мкм). Для возбуждения потенциала действия подавалось деполяризирующее напряжение (полярность напряжения противоположна полярности потенциала покоя).
В допороговой области (Io ≤ Ic) данное воздействие приводило лишь к смещению исходного отрицательного потенциала. При превышении порога – наблюдалась генерация потенциала действия - быстрая деполяризация мембраны (изменение знака потенциала) с последующей реполяризацией. Зависимость потенциала действия от времени имела сложный и немонотонный характер. Потенциалы, регистрируемые в различных точках имели один и тот же вид, но были сдвинуты по времени
Измерения показали, что пороговый ток возбуждения Ic существенно зависит от длительности импульса. При очень больших длительностях этот ток минимален, но не равен нулю. Это минимальное значение тока Imin называется реобазой. При уменьшении длительности импульсов значение порогового тока возрасатает, причем в области малых длительностей порог постоянен по заряду (qmin =Ic* Δt = const) , а не по току. Отношение порогового заряда к реобазе
Δtхр = qmin/ Imin соответствует переходу от одного типа поведения потенциала к другому типу поведения.
Если сообщить аксону два последовательных импульса возбуждения, то его отклик зависит от интервала времени, разделяющего эти импульсы. При малом интервале времени второй импульс приходит в момент времени, в которой аксон еще возбужден предыдущим импульсом, т.е. находится в абсолютном рефрактерном состоянии и не может возбудиться снова. Затем наступает относительное рефрактерное состояние, когда для возбуждения аксона требуются большие амплитуды импульса. Продолжительность рефрактерного периода – несколько миллисекунд.
Приведенные экспериментальные данные свидетельствуют о сложных механизмах, лежащих в основе распространения нервного импульса. Действительно, прямое кабельное распространение невозможно из-за высокого электрического сопротивления аксоплазмы .
Задача:Рассчитать сопротивление участка гигантского аксона кальмара с диаметром 600мкм и длиной 1м используя максимальные значения удельной электропроводности аксоплазмы – 10 Ом.м.
Ответ: около 300 Мом.
При столь высоких сопротивлениях электрический сигнал должен сильно затухать, а экспериментально было установлено, что потенциал действия распространяется практически без затухания.
Действительный механизм распространении нервного импульса был предложен Ходжкиным и Хаксли. Они обратили внимание на то, что при удалении из внеклеточной среды ионов натрия потенциал действия не возникает. Таким образом, его появление было объяснено резким возрастанием проводимости мембраны относительно ионов натрия. Это подтверждается и тем, что на пике потенциала действия его значение близко к равновесному значению для ионов натрия.
Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na+ , K+, Са2+ и Сl-. Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (илиэлектровозбудимые), например быстрые Na+ каналы, и лиганд-управляемые (или хемовозбудимые). Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами, белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — "закрыт» —и т.д.).
Дата добавления: 2018-05-10; просмотров: 867;
Поиск по сайту
Узнать еще
- Appeal –привлекательность.
- Applications (приложения)
- Cила резания при точении
- Cущность организации и ее основные признаки
- D-технология построения чертежа. Типовые объемные тела: призма, цилиндр, конус, сфера, тор, клин. Построение тел выдавливанием и вращением. Разрезы, сечения.
- D-элементы, их применение в медицине и фармации.
- F50 Расстройства приема пищи
- He рекомендуем использовать данный метод, если в дальнейшем будет необходимость прибегнуть к отгибу приборной панели.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
