ТИПЫ МЕЖПОПУЛЯЦИОННЫХ ВЗАИМОДЕЙСТВИЙ
Организмы популяций разных видов могут либо влиять, либо не влиять друг на друга. Влияние, в свою очередь, делится на благоприятное или неблагоприятное.
Удобную и наглядную форму оценки влияния разного рода предложил Ю. Одум (1975). Он обозначил благоприятное влияние знаком «+», неблагоприятное «–» и нейтральное 0. Например, если ни одна из двух популяций не влияет на другую, то взаимодействие обозначают (0 0); при обоюдных вредных влияниях - (– –), при взаимовыгодной связи — (+ +). Другие возможные связи обозначаются как (+ –), (– 0), (+ 0). Существует шесть вариантов взаимодействий (табл. 1).
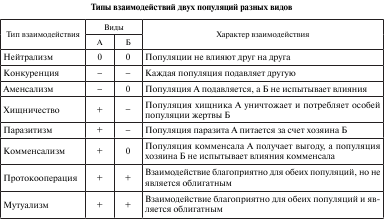
1.1. Конкурентные взаимоотношения
Организмы растут, размножаются, расселяются, гибнут. Они подвержены влиянию условий среды, в которой живут, и ресурсов, которые они потребляют. Не существует организмов, живущих в полной изоляции. Любой организм входит в состав популяции, включающей особей того же вида. Особи одного вида имеют наиболее сходные потребности, чем особи разных видов. И те, и другие должны удовлетворить свои потребности в необходимых ресурсах, чтобы выжить, вырасти и оставить потомство.
Однако их суммарные потребности в каком-либо ресурсе могут превышать его запас. В таком случае между особями возникает конкуренция за ресурс. Конкуренция – взаимодействие особей, вызванное сходными потребностями в ограниченном ресурсе, которое приводит к снижению выживаемости, скорости роста и (или) размножения конкурирующих особей.
Истощая или используя часть находящегося в недостатке ресурса, конкуренты могут прямым путем уменьшать его количество, доступное другим организмам. Конкуренция путем прямого влияния, например агрессивных столкновений или выделения токсических веществ, подавляющих конкурента, называется интерференционной. Этот вид конкуренции часто встречается среди подвижных животных, например волков, медведей, защищающих свою территорию, в результате чего сама территория принимает статус ресурса. Примером этого вида конкуренции является также прямая борьба между самцами за право оставить потомство. Массовое развитие цианобактерий и переход их в статус доминантной группы основаны на выделении ими токсических веществ в воду и подавлении развития планктонных водорослей других таксонов.
Косвенное подавляющее влияние, возникающее в результате уменьшения доступности общего ресурса, известно как эксплуатационная конкуренция.
Результат отрицательного воздействия конкурирующих сторон может быть обоюдоравным, либо один из конкурентов оказывает большее отрицательное воздействие на другой. Первый вариант конкуренции рассматривают как симметричную, второй – как асимметричную. Итоговый результат для двух вариантов конкуренции будет разным. В первом случае обоим конкурентам придется поделить на равные части ограниченный ресурс, во втором — более сильный конкурент способен потеснить либо полностью подавить и вытеснить более слабого.
Поскольку для каждой из взаимодействующих сторон конкуренция сказывается отрицательно, а избегание конкуренции всегда выгодно, то она является мощным эволюционным фактором, приводяшим путем дивергенции к разделению ниш, специализации видов и возникновению биоразнообразия.
Межвидовая конкуренция основана на использовании ограниченных в запасе сходных ресурсов популяциями разных видов. И действительно, для роста и жизнедеятельности растений необходим практически одинаковый набор основных биогенных, макро- и микроэлементов, однако в разных сочетаниях и количествах с учетом их стадийного развития. Большинство животных олиго-, поли-, пантофаги, но не монофаги. Следовательно, общность потребляемых ресурсов является основой для формирования у них межвидовых взаимодействий. При ограниченности запасов общих ресурсов естественное следствие – межвидовая конкуренция.
Примеров наличия в природе межвидовой конкуренции в научной литературе достаточно. Почти 100 лет назад один из известных «отцов-основателей» экологии растений А. Г. Тенсли изучал конкуренцию между двумя видами подмаренника. Galium hercinicum растет в Великобритании на кислых почвах, тогда как распространение Galium pumilum ограничено щелочными почвами. Выращивая виды по отдельности, А. Г. Тенсли обнаружил, что каждый из них хорошо рос как на кислой, так и на щелочной почве, взятой из природных местообитаний каждого из них. Однако при совместном выращивании на кислой почве успешно рос только G. hercinicum, а на щелочной – только G. рumilum. В данном примере исход конкуренции зависел от условий произрастания растений разных видов.
Сходный результат наблюдается также в том случае, если одна из популяций вырабатывает химическое вещество, действие которого подавляет или ограничивает развитие конкурирующей с ней популяции другого вида. Такие химические воздействия называют аллелопатией (от греч. аllelon – друг друга и pathos – страдание, болезнь).
Так, разлагающиеся листья каштанов выделяют в почву токсическое вещество, подавляющее рост сеянцев многих других видов деревьев. Лещина и дуб своими выделениями часто угнетают рост травянистых растений под кроной. Такое взаимодействие можно рассматривать как крайнюю форму проявления асимметричной конкуренции – аменсализма.
Подробно исследованы аллелопатические ингибиторы, выделяемые колючими кустарниками, образующими сообщество, которое в Калифорнии называется чапарраль. Оно представляет собой чередующиеся купы кустарников и лугов. Вокруг многих кустарников чапаррали существуют многометровые «пролысины» — кольца отчуждения, где не растет травянистая растительность, поскольку в почве накапливаются токсины группы терпенов. Примерно четыре раза за столетие чапарраль выгорает и пламя пожара разрушает терпены, после чего весь участок зеленеет буйными лугами. Однако с появлением кустарников «пролысины» восстанавливаются.
Были исследованы химическая природа ингибиторов и их физиологическое действие, что позволило выявить их важную роль в регуляции видового состава сообщества чапаррали и его динамики. Так, летучие токсины, образуемые в листьях (особенно цинеол и камфора) двух видов ароматических кустарников, подавляют рост травянистой растительности. В сухой период листья накапливаются в почве в таком количестве, что с наступлением сезона дождей они подавляют прорастание семян и рост растений в обширной зоне вокруг каждой группы кустов. Другие виды кустарников образуют водорастворимые фенолы и алкалоиды, которые также дают им занять доминирующее положение в сообществе чапаррали. Однако периодические пожары, уничтожая источник токсинов и очищая от них почву, приводят к тому, что здесь появляются растения, семена которых не погибли во время пожара. В результате в дождливый сезон, следующий за пожаром, наблюдается активный рост однолетних растений, который продолжается затем каждую весну до тех пор, пока в этом месте снова не появятся кустарники и их токсическое действие не достигнет достаточной эффективности. Таким образом, за счет попеременного действия пожаров и токсинов происходят циклические изменения видового состава, что является адаптивной особенностью экосистемы этого типа.
Р. Уиттэкер (1970) в своем обзоре растительных ингибиторов приходит к заключению о том, что высшие растения синтезируют значительное число веществ, являющихся репеллентами или ингибиторами для других организмов. Аллелопатия оказывает существенное влияние на скорость растительных сукцессий, а также на видовой состав растительных сообществ. Химические воздействия двояко сказываются на разнообразии видов в природных сообществах: в некоторых сообществах, где доминирующее положение занимает какой-то один вид, характеризующийся интенсивным аллелопатическим влиянием, разнообразие видов невелико, в то же время в других сообществах благодаря различным химическим приспособлениям разнообразие видов оказывается значительным.
Описанные выше способы борьбы с конкурентами действуют в природных условиях одновременно, и поэтому оценить роль каждого из них в многовидовом сообществе практически невозможно. На помощь приходят лабораторные эксперименты с участием двух-трех взамодействующих видовых популяций, позволяющие выявить закономерности конкуренции.
Классическим примером изучения межвидовой конкуренции являются лабораторные эксперименты русского эколога Г. Ф. Гаузе (1934, 1935) на трех видах инфузорий рода Paramecium. Популяции всех трех видов хорошо росли в монокультуре, достигая в пробирках с жидкой средой стабильных значений предельных плотностей. Пищей инфузориям служили бактериальные или дрожжевые клетки, растущие на регулярно добавляемой овсяной муке.
При совместном выращивании P. сaudatum и P. аurelia численность P. caudatum всегда снижалась вплоть до полного вымирания.
Однако при совместном выращивании P. аurelia и P. bursaria, наблюдалось сосуществование, но на уровне более низких величин плотностей, чем в монокультурах, что свидетельствует о наличии межвидовой конкуренции эксплуатационного типа. Как выяснилось, выживанию каждой из них помогло пространственное разобщение в пробирке: P. аurelia держалась в толще культуральной среды и питалась бактериями, тогда как P. bursaria концентрировалась у дна пробирок, потребляя дрожжевые клетки.
При совместном выращивании P. caudatum и P. аurelia происходило полное подавление (исключение) популяции первого вида вторым. В случае с совместным выращиванием P. аurelia и P. bursaria оба конкурента сосуществовали, так как отличались трофической и пространственной составляющими своих ниш. Следовательно, их сосуществование было связано с дифференцировкой реализованных ниш или «разделением ресурсов».
Результаты экспериментов Г. Ф. Гаузе дают замечательную иллюстрацию результатов межвидовой конкуренции с точки зрения теории экологической ниши. Вспомним, что в отсутствие конкурентов другого вида (т. е. при выращивании парамеций в монокультуре) экологическая ниша представляет собой фундаментальную нишу. Однако при совместном культивировании разных видов (пример с P. caudatum и P. аurelia) приходится довольствоваться реализованной нишей. В результате недостатка ресурсов для более слабого конкурента реализованная ниша становится столь малой, что он не в состоянии далее жить и успешно размножаться.
Полученные результаты по изучению межвидовой конкуренции в разных условиях позволили сформулировать принцип конкурентного исключения, впоследствии названный принципом Гаузе: если два конкурирующих вида сосуществуют в стабильных условиях, то это происходит благодаря дифференциации ниш, т. е. разделения реализованных ниш этих видов; если, однако, такой дифференциации не происходит или если ей препятствуют условия среды, то один из конкурирующих видов будет истреблен или вытеснен другим; такое вытеснение наблюдается тогда, когда реализованная ниша более сильного конкурента полностью перекрывает те части фундаментальной ниши более слабого конкурента, которые имеются в данном местообитании.
Принцип Гаузе получил широкое признание благодаря многочисленности подтверждающих его фактов, а также теоретическим предпосылкам, положенным в математическую модель Лотки – Вольтерры, позволяющую определить наличие и силу конкуренции между популяциями разных видов.
Модель межвидовой конкуренции Лотки – Вольтерры построена на использовании логистического уравнения роста популяции для определения исхода конкурентного взаимодействия. Логистическое уравнение 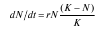 содержит в скобках выражение, отражающее существование внутривидовой конкуренции. В основе модели Лотки – Вольтерры лежит замена этого выражения таким, которое отражает как внутривидовую, так и межвидовую конкуренцию.
содержит в скобках выражение, отражающее существование внутривидовой конкуренции. В основе модели Лотки – Вольтерры лежит замена этого выражения таким, которое отражает как внутривидовую, так и межвидовую конкуренцию.
Численность популяции вида 1 обозначена N1, а численность вида 2 –N2. Предельная плотность насыщения и максимальная врожденная скорость роста популяций обозначены K1, K2, r1 и r2 соответственно.
На модели Лотки – Вольтерры, описывающей межвидовую конкуренцию, можно получить несколько конечных результатов: безусловное исключение одного вида другим, исключение в зависимости от соотношения величин начальной плотности и устойчивое сосуществование. Следует подчеркнуть, что в ходе проведения экспериментальных исследований конкуренции все остальные условия были постоянными.
В отличие от эксперимента, в природе все сложнее, так как в ней происходит одновременное внутривидовое и межвидовое взаимодействие, в котором участвуют популяции не двух, а более видов. Кроме того, в природный среде на итог конкуренции накладывается влияние непредсказуемых изменчивых условий разного рода, в том числе и климатических. В флуктуирующей среде равновесие между конкурирующими видами неоднократно нарушается и преимущество будет переходить от одного вида к другому. Следовательно, сосуществование возможно просто благодаря изменению условий среды. Такой аргумент использовал Дж. Хатчинсон для объяснения «планктонного парадокса» – явления сосуществования многочисленных видов планктонных организмов в простой среде, где, по-видимому, мало возможностей для разделения ниш.
Дж. Хатчинсон предположил, что среда, хотя она и очень проста, постоянно претерпевает различные изменения, в частности сезонные. В любой отдельный промежуток времени условия среды могут способствовать вытеснению определенного вида, однако условия меняются, и еще до того, как данный вид окажется окончательно вытесненным, они могут сложиться благоприятно для его сосуществования. Другими словами, исход конкурентных взаимодействий в равновесном состоянии может не играть решающей роли, если условия среды изменяются задолго до того, как достигается равновесие. А поскольку любая среда изменчива, равновесие между конкурентами должно постоянно сдвигаться и сосуществование будет наблюдаться при таком разделении ниш, при котором в стабильных условиях происходило бы исключение одного из видов. Имеются примеры, показывающие возможность сосуществования сильного и слабого конкурентов в изменчивых условиях эфемерных биотопов.
Пример такого сосуществования двух видов легочных моллюсков в мелководных прудах приведен в работе В. Брауна (Р. Бигон, Дж. Харпер, К. Таунсенд, 1989). При искусственном изменении плотности одного или другого вида в естественных условиях пруда наблюдалось снижение плодовитости Physa gyrina под влиянием конкуренции со стороны Lymnaea elodes, но такое влияние не было обоюдным. Если конкуренция продолжалась в течение всего лета, то более сильным конкурентом оказывался L. elodes. Однако P. gyrina начинал размножаться раньше, достигая меньших размеров, чем P. elodes, и во многих прудах, которые к началу июля пересыхали, покоящиеся яйца часто оставлял только один из видов. Несмотря на то, что более слабым конкурентом бесспорно была P. gyrina, в целом по прудовым биотопам наблюдалось сосуществование этих видов. К сосуществованию сильного и более слабого конкурентов в одном биотопе приводит также характер пространственного распределения. При групповом размещении более сильного конкурента свободные от него участки в местообитании занимаются популяцией слабого конкурента. При равномерном распределении у более сильного конкурента больше шансов вытеснить слабого.
В средах с большой пространственной неоднородностью отмечается более высокое видовое богатство из-за наличия в них большего разнообразия микроместообитаний, более широкого диапазона микроклиматических условий, больших возможностей укрытия от хищников. Другими словами, гетерогенность условий приводит к расширению спектра ресурсов, что может внести свои коррективы в итог межвидовой конкуренции.
Самым важным следствием межвидовой конкуренции, а также отличием от внутривидовой, является ее влияние на видовую, трофическую структуру биоценоза, а также пространственное распределение особей по биотопу.
1.2. Взаимоотношения хищник жертваª
Взаимоотношения хищник – жертва включают все взаимодействия, при которых одни организмы – хищники – используют в пищу другие – жертв, причем жертва должна быть живой перед первым нападением на нее хищника. Такое определение исключает детритофагию, или потребление мертвого органического вещества.
Хищничество представляет собой главную силу, обеспечивающую передвижение вещества и энергии в биоценозах.
Существует два основных подхода к классификации хищников. Наиболее естественной и привычной является таксономическая основа классификации: хищники в собственном значении этого слова нападают и поедают живых животных, растительноядные организмы – растения, а всеядные – и тех, и других.
Согласно другому подходу, предложенному Д. Томпсоном (1982), хищником является организм, поедающий живую жертву независимо от ее таксономичесой принадлежности, т. е. как животного, так и растительного происхождения. Поэтому понятие хищника в этой трактовке более широкое. Согласно данной классификации, названной функциональной, выделяют четыре основных типа хищников: истинные хищники, хищники с пастбищным типом питания, паразитоиды и паразиты. Авторы полагают, что выделение различных типов хищников позволяет более полно понять сущность хищничества, поскольку конечный результат от нападения хищников разных типов различен.
Истинные хищники убивают свою жертву сразу после нападения и поедают ее целиком либо только ее часть. В течение всей жизни они убивают много разных особей жертвы. К этому типу хищников относятся тигры, львы, божьи коровки, насекомоядные растения, муравьи, фильтрующие планктон киты.
Хищники с пастбищным типом питания в течение своей жизни также используют большое число жертв, но, как правило, съедают только часть жертвы. Их воздействие на жертву может быть различным, но чаще всего вредным. Нападение такого хищника редко приводит к скорой гибели жертвы. Примерами хищников этого типа являются травоядные позвоночные, мухи, кусающие позвоночных животных, пиявки, сосущие у них кровь.
Паразиты, как и хищники с пастбищным типом питания, обычно поедают только часть своей жертвы (хозяина). Их нападение на жертву причиняет ей вред, но редко приводит к летальному исходу. В отличие от хищников с пастбищным типом питания, паразиты в течение своей жизни нападают только на одну или немногих особей жертвы. Они теснее связаны с жертвой, что не характерно для хищников двух предыдущих типов. Примеры паразитов всем известны: ленточные черви, печеночный сосальщик, туберкулезная палочка, вирусы кори и табачной мозаики, ржавчинные грибы, головня, омела белая и др.
Паразитоиды – группа насекомых отряда Перепончатокрылые и некоторых видов отряда Двукрылые, взрослые особи которых ведут свободный образ жизни, а яйца откладывают в тело другого насекомого. Паразитоиды тесно связаны с определенным организмом хозяина. Вылупившиеся из яиц личинки паразитоидов развиваются внутри (реже на поверхности) своего хозяина, приводя его к гибели. Примерами паразитоидов являются наездник трихограмма, его самка заражает яйца бабочек (например, яблонной плодожорки), а также бычий овод, яйца которого самка откладывает под кожу крупного рогатого скота, что приводит к истощению животных.
Последствия хищничества. Истинные хищники оказывают летальное воздействие на отдельную особь жертвы. Однако на популяцию жертвы в целом эффект влияния будет другой, поскольку уничтоженные особи не представляют собой случайную выборку из всей популяции, а избежавшие гибели особи часто проявляют реакции, направленные на компенсацию потери популяции. Исследования свидетельствуют, что наиболее вероятными жертвами становятся молодые, бездомные, больные, травмированные и одряхлевшие животные, т. е. легкая для поимки добыча. Поэтому роль хищников рассматривают как санитарную, проявляющуюся в снижении угрозы распространения заболеваний, элиминации неполноценных особей и поддержании популяции жертвы в здоровом состоянии.
Сходные результаты получены и для популяций растений. Так, в Австралии гибель зрелых эвкалиптов, вызванная уничтожением листьев пилильщиками (Perga affinis affinis), почти полностью ограничивалась ослабленными деревьями, произраставшими на бедных почвах, или деревьями, пострадавшими от повреждения корней (М. Бигон, Дж. Харпер, К. Таунсенд, 1989).
Влияние растительноядности на растение зависит от того, в каком периоде жизненного цикла находятся растения и какие именно его части подвергаются поеданию. Откусывание листьев, высасывание сока, минирование, поедание меристемы, повреждение цветков или плодов, подгрызание корней – все это по-разному влияет на растение. Кроме того, поскольку растение после воздействия хищника остается некоторое время живым, эффект этого воздействия в значительной степени зависит от реакции самого растения. Так, может перераспределиться поток минеральных солей и органических веществ из одной части растения в другую, или измениться уровень обмена веществ, относительная скорость роста корней, могут пробудиться спящие почки, образоваться защитные вещества и ткани. От скорости реализации этих реакций будет зависеть степень повреждающего воздействия хищника на растение.
Хищничество – сложный процесс, состоящий, как правило, из серии последовательных событий: обнаружение жертвы, нападение на нее, поимка, поедание. На эти процессы накладываются влияния особенностей поведения как хищника, так и его жертвы, а также изменчивость экологической ситуации. Для хищников необходимо некоторое минимальное количество пищи, обеспечивающее поддержание жизнедеятельности организма, которое называется пороговым. Рост и размножение популяции происходит только при концентрации пищи выше пороговой. На разных видах хищных животных обнаружена четко выраженная закономерность: с увеличением плотности жертвы скорость потребления пищи возрастает, но постепенно этот рост замедляется и, наконец, приостанавливается независимо от повышения плотности жертвы. Концентрация пищи жертвы, при которой значения скоростей роста хищника максимальны, называется насыщающей.
Степень воздействия хищников на своих жертв во многом определяется наличием или отсутствием предпочтения к определенному виду жертвы – избирательностью питания.
Один из методов оценки избирательности питания предложил В. С. Ивлев (1977).
Пищевое предпочтение определяется качеством и количеством пищи, размером и доступностью жертвы. Среди хищников встречаются монофаги (одноядные), олигофаги (ограниченноядные), полифаги (многоядные) и пантофаги (всеядные). Хищничество имеет широкий спектр различий в рационе: от питания одним видом жертвы – монофагии – до всеядности – пантофагии.
Эволюционные процессы, приводящие к ограничению рациона – монофагии, вызваны тем, что жизнедеятельность хищника тесно связана с жизнедеятельностью его жертвы или хозяина, а также когда отдельная особь хищника имеет дело с одной жертвой. Пока какой-либо вид жертвы остается многочисленным, доступным и предсказуемым, отбор будет благоприятствовать все большей специализации, ведущей к монофагии. Такая специализация повышает эффективность питания видов-специалистов и, по крайней мере, частично позволяет им уйти от межвидовой конкуренции.
Вместе с тем полифагия также дает определенные преимущества. Хищник может составить себе сбалансированную диету, подбирая ее из ряда различных кормовых объектов, и способен поддерживать этот баланс, меняя предпочтение в зависимости от конкретных условий. Кроме того, полифаг легко находит корм, затраты на поиск обычно невелики, и отдельные особи редко голодают из-за того, что обилие одного из видов пищи колеблется. Нередки случаи переключения с одного вида жертвы на другой в связи с изменением соотношения численности популяций, предпочитаемых по размеру и пищевым качествам. Например, у гладыша обыкновенного Notonecta glauca переключение происходит потому, что он приобретает опыт в нападении на пресноводных равноногих ракообразных по мере того, как они становятся многочисленнее в сравнении с альтернативной жертвой – личинками поденок.
Животные, у которых полностью или частично отсутствует избирательность в потреблении пищевых объектов, называются неспециализированными хищниками или генералистами. К ним относится большинство хищников с пастбищным типом питания.
И действительно, крупный рогатый скот, кролики имеют низкую избирательность питания. При достаточно высокой численности таких хищников они способны держать травостой в состоянии низкорослого дерна. Следствием такого безвыборочного питания будет поддержание высокого видового богатства растительности благодаря подавлению межвидовой конкуренции у жертвы. Так, исследования показали, что многолетняя защита травостоя от поедания путем огораживания приводит к гибели 45 % видов растений, вытесненных оставшимися более сильными конкурентами.
Кроме поддержания высокого флористического разнообразия, умеренное потребление растительности приводит к усилению интенсивности фотосинтеза за счет уменьшения самозатенения растений, что повышает продуктивность растений жертв.
Однако значительная плотность хищников с пастбищным типом питания, потребляющих не только надземные части растения, но и подземные (корни), как это типично для коз, приводит к полному истреблению растений жертв.
Специализированные хищники (специалисты) питаются одним или немногими видами жертв. Как уже указывалось выше, они, в целях экономии энергии, нападают на более доступную добычу, выполняя таким образом санитарную функцию. Поскольку наличие пищевой специализации ограничивает возможность переключения на другие виды пищи, то следствием является возникновение взаимосвязанных циклических колебаний численности хищника и жертвы. Циклические колебания численности с периодичностью 10 –11 лет наблюдаются у канадской рыси и ее жертвы – зайца беляка. При низкой численности как хищника, так и жертвы численность зайца возрастает из-за его большей плодовитости, быстрого созревания и, как следствие, большей скорости роста популяции, чем у рыси.
По мере возрастания популяции зайца начинает увеличиваться и популяция хищника, но с некоторым отставанием. Наконец, отлов жертвы хищником становится чрезмерным и численность жертвы начинает сокращаться. Низкая численность жертвы приводит к сокращению численности хищника, так как отыскание и поимка жертвы энергетически становится невыгодной. Затем цикл вновь повторяется. Так устанавливается динамическое равновесие между хищником и жертвой, что вносит свой вклад в поддержание стабильности биоценоза.
Стратегия хищника в добывании жертвы состоит в увеличении эффективности питания, т. е. в большей энергетической выгоде от съеденной пищи по сравнению с ее затратой на поиск пищи и ее потребление. В этом плане интересен пример взаимодействия популяций волков и лосей в провинции Квебек (Канада) (А. М. Гиляров, 1990). Оказалось, что волки не заселяют местность при численности лосей менее 0,2 особей/км2. При увеличении плотности лосей от 0,2 до 0,5 особей/км2 пресс волков значительно возрастает. Дальнейший рост плотности лосей (свыше 0,5 особей/км2) не приводит к возрастанию плотности волков.
Взаимные приспособления хищника и жертвы. В ходе длительной взаимной связи хищника и жертвы у первого вырабатываются эффективные средства нападения, у жертвы — эффективные средства защиты. Подобные эволюционные изменения двух или более взаимодействующих видов называются коэволюцией.
На долю наземных растений приходится большая часть массы всего живого вещества биосферы. Однако только около 7 % продукции живой массы растений служит фитофагами в пастбищных цепях питания, остальная масса направляется в детритные цепи. Низкий процент использования живой массы растений хищниками с пастбищным типом питания свидетельствует о наличии у них эффективных способов защиты от выедания.
Защита растений от истребления фитофагами идет по пути образования защитных морфологических структур (шипы, колючки, толстая кора и т. д.), а также за счет выработки ими средств химической защиты, делающих ткани растений несъедобными или даже ядовитыми. Среди этих веществ есть алкалоиды (например, никотин в листьях табака), таннины, различные смолы, стероиды и др. Большая часть этих веществ не просто вредные продукты метаболизма, которые потом выводятся растениями при листопаде, они специально синтезируются растением. Так, у астрагала вырабатывается мизеротоксин, вызывающий паралич дыхания и даже гибель овец и крупного рогатого скота. Ядовиты зверобой продырявленный, вех ядовитый и др.
Некоторые растения приобрели способность синтезировать биологически активные вещества, обладающие высочайшей специфичностью действия только по отношению к определенному виду потребителя. Примером может служить бальзамическая пихта (Abies balsamea), которая синтезирует вещество ювабион, являющееся аналогом ювенильного гормона насекомых. В ходе нормального развития насекомых прекращение синтеза ювенильного гормона приводит к тому, что активными становятся другие гормоны, способствующие нормальному завершению метаморфоза. Если же в организм насекомого продолжает поступать ювенильный гормон (в данном случае ювабион), то дополнительно происходит еще 1-2 линьки, после чего личинка погибает.
Таким образом, синтезируя ювабион, бальзамическая пихта защищает свои листья от истребления личинками насекомых.
Защитные приспособления животных от хищников разнообразны. Это и покровительственная окраска, и особая форма тела, делающие их трудно различимыми в среде обитания, быстрая скорость передвижения и маневренность, помогающие жертве уйти от преследования хищника. У многих животных, не имеющих укрытий, своеобразным способом компенсации потерь от хищников является интенсификация размножения.
Этот способ развился, например, у тлей, планктонных ракообразных и коловраток, приобретших партеногенетический способ размножения.
В качестве средства защиты от поедания многие животные, как и растения, вырабатывают яды и токсины. Так, некоторые виды многоножек в качестве оборонительного средства используют синильную кислоту, вызывающую у нападающих на них животных судороги, удушье, паралич. Жуки нарывники и шпанские мушки продуцируют кантаридин – вещество кожно-нарывного действия. Батрахотоксин ядовитых лягушек Колумбии нарушает у употребивших их в пищу хищников нормальную работу натриевых каналов нервных клеток и мышечных волокон, что приводит к деполяризации мембран. Токсин европейских видов жаб – буфоталин –останавливает у хищника работу сердца в систоле.
У паразитов развились приспособления, позволяющие потреблять пищу хозяина, не приводя его к гибели. К такого рода приспособлениям относятся смена хозяина в жизненном цикле паразита, сезонность заражения. Адаптации хозяина (жертвы) направлены на повышение иммунитета и устойчивости к воздействию паразита. В ходе эволюции организмы не могут эффективно специализироваться сразу по нескольким направлениям. Например, за высокую степень защищенности от хищников и высокую конкурентоспособность приходится расплачиваться сокращением скорости индивидуального и популяционного роста. Так, например, у ели в благоприятный для размножения семенной год (большая продукция семян) прирост уменьшается на 38 % по сравнению с другими годами, а прирост годовых колец у лесных деревьев в урожайный на семена год может уменьшиться так же сильно, как при полном объедании листьев гусеницами. Годы с низким урожаем семян, таким образом, являются периодами, в которые происходит восстановление затраченных на размножение ресурсов дерева. Эти примеры показывают, что невозможно найти организм, способный одновременно быть r и K-стратегом.
Основываясь на этих соображениях, Р. Кейтс и Г. Орианс предположили, а потом и доказали в экспериментах на слизнях, что растения поздних стадий сукцессий лучше защищены от фитофагов, чем раннесукцессионные, которым свойственно быстро размножаться, но они не способны вынести пресса конкурентов и фитофагов.
Видовые популяции хищников и жертв, являясь взаимосвязанными эволюционирующими единицами, вырабатывают сложные механизмы приспособлений, способствующих в конечном счете сохранению системы «хищник – жертва».
Исследования взаимоотношений «хищник – жертва» в природных и лабораторных условиях позволяют определить долю изъятия из популяций жертвы, при которой в ней будет сохраняться оптимальный и устойчивый выход продукции. Одна из экспериментальных моделей, аквариумную рыбку гуппи (Lebistes reticulates) использовали для имитации популяции промысловой рыбы. Можно видеть, что максимальный устойчивый выход продукции получали в том случае, когда в каждый репродуктивный период изымалась одна треть популяции, что приводило
к уменьшению равновесной плотности до величины, составляющей меньше половины плотности не облавливаемой популяции. Эта 30 %-я оптимальная величина изъятия характерна для популяции гуппи. Для популяции другого вида рыб с меньшим биотическим потенциалом, чем у гуппи, следует ожидать меньшей оптимальной величины изъятия. Таким образом, связь величины изъятия без подрыва вклада продукции эксплуатируемой популяции в природную экосистему напрямую зависит от характерного для нее биотического потенциала.
Краткое рассмотрение взаимоотношений хищника и жертвы показывает, что они разнообразны и сложны. Представленные примеры приводят к выводу о важной роли хищничества в популяционных процессах всех видов. Если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне, который ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы.
Хищничество способно нарушить очень тонкое равновесие между популяциями разных видов, изменить структуру звеньев многовидового сообщества. Особенно опасен новый хищник, который может сдвинуть это равновесие в пользу одного или другого вида. Рассмотрим пример влияния нового хищника на перестройку всей трофической структуры водной экосистемы, приведенный в учебнике Р. Риклефса «Основы общей экологии» (1979).
Гатун – искусственное озеро, образовавшееся в начале прошлого века при постройке Панамского канала. За несколько десятков лет жизни озера в нем сформировалась стабильная экосистема, состоящая из фитопланктона, зоопланктона и планктоноядной рыбы. Рыба удерживала численность зоопланктона на низком уровне. В отсутствии обильного зоопланктона плотность популяций разных видов фитопланктона возрастала до такого уровня, что озерная вода имела вид зеленого супа («цветение воды»). В 1967 г. в озеро Гатун была случайно интродуцирована рыба туканоре – один из видов цихлид, которая оказалась прожорливым и активным хищником, поедающим другие виды рыб. Основная пищевая цепь озера изменилась и стала содержать не три, а четыре звена: фитопланктон, зоопланктон, планктоноядные рыбы и хищная рыба туканоре. Размножившись, новый вселенный хищник понизил численность популяций планктоноядных местных рыб, что привело к увеличению популяций зоопланктона, а это, в свою очередь, повлекло за собой заметное снижение популяций фитопланктона. Поэтому вода в озере стала более прозрачной. Как видно, вселение одного хищника оказало глубокое влияние на обилие и продуктивность трех нижележащих трофических уровней. Кроме того, в результате конкуренции с туканоре сократились популяции таких птиц, питающихся рыбой, как крачки и зимородки, а популяции комаров, водными личинками которых обычно питаются мелкие рыбы, увеличились. Таким образом, вселение хищной рыбы серьезно изменило трофическую структуру не только озера, но и околоводных животных.
1.3. Взаимоотношения паразит хозяин
Паразиты и хищни<
| <== предыдущая лекция | | | следующая лекция ==> |
| | | Понятие государственного органа |
Дата добавления: 2022-04-12; просмотров: 160;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
