Соотношение понятий «биогеоценоз» и «экосистема»
Единство биоценоза и заселяемого им биотопа образует экологическую систему (см. рис. 1). Экологическая система, или экосистема, – пространственно определенная совокупность живых организмов разных таксонов (биоценоз) и среды их обитания (биотоп), объединенных вещественно-энергетическими и информационными взаимодействиями.
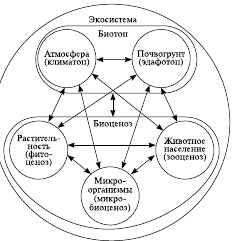
Термин «экосистема» ввел английский ботаник А. Тенсли (1935). Понятие экосистемы не ограничивается какими-то признаками ранга, размера, сложности или происхождения. Поэтому оно приложимо как к относительно простым искусственным (аквариум, теплица, пшеничное поле, обитаемый космический корабль), так и сложным естественным комплексам организмов и среды их обитания (озеро, лес, океан, река, пруд).
Параллельно с развитием понятия «экосистема» в первой половине ХХ в. развивалось учение о биогеоценозах (греч. bios – жизнь, ge –земля, koinos – общий). Этот термин предложил русский ученый академик В. Н. Сукачев в 1940 г. Для большинства биогеоценозов основной характеристикой является то, что его протяженность четко определяется границами наземного растительного сообщества –фитоценоза. Например, легко различимы между собой биогеоценозы соснового, елового или широколиственного лесов. Там, где нет четко выраженного фитоценоза, нет и биогеоценоза.
Понятия «экосистема» и «биогеоценоз» тождественны только для природных наземных образований, четко очерченных границами фитоценоза. Если выделить границы фитоценоза сложно, то применяется понятие «экосистема». Поэтому любой биогеоценоз является экосистемой, но не любая экосистема будет биогеоценозом. Таким образом, экосистема – понятие более общее, а биогеоценоз является частным случаем экосистемы. Поэтому именно экосистемы рассматриваются в качестве слагающих биосферу «кирпичиков», ее миниатюрных модулей. Совокупность всех экосистем нашей планеты создает гигантскую экосистему, называемую биосферой.
2.Пищевые цепи и пищевые сети
В биоценозах существуют сложные пищевые взаимодействия между автотрофными и гетеротрофными организмами. Одни организмы поедаются другими, благодаря чему формируется цепной процесс переноса вещества и поток энергии в экосистемах, лежащий в основе круговорота веществ в природе.
Перенос вещества и энергии от ее источника – автотрофов – к потребителям – гетеротрофам, происходящий в результате последовательного поедания одними организмами других, называется пищевой цепью. Отдельное звено пищевой цепи является трофическим уровнем. Как установлено Р. Линдеманом (1942), при каждом очередном переносе большая часть (80-90 %) потенциальной энергии теряется на дыхание, тепло и др. Это ограничивает возможное число этапов, или звеньев, пищевой цепи, как правило, до четырех-пяти. Пример обычной пищевой цепи в пресноводном водоеме, состоящей из пяти трофических уровней: организмы фитопланктона (первый трофический уровень – продуценты) – рачки дафнии (второй трофический уровень – консумент I порядка) – рыба плотва (третий трофический уровень – консумент II порядка, или хищник первого порядка) – щука или судак (четвертый трофический уровень – консумент III порядка, или хищник второго порядка), выдра (пятый трофический уровень – консумент IV порядка, или хищник третьего порядка). Следует подчеркнуть, что приведенная трофическая классификация делит на трофические уровни не сами виды, а их типы жизнедеятельности. Популяция одного вида может принадлежать к одному или более трофическим уровням в зависимости от того, какие источники энергии она использует.
Трофическую структуру экосистемы можно изобразить графически в виде экологической пирамиды, основанием которой служит первый уровень продуцентов, а остальные уровни консументов образуют последующие этажи пирамиды. Существует три способа построения экологической пирамиды: 1) пирамида чисел, отражающая численность отдельных организмов; 2) пирамида биомассы, характеризующая общий сухой вес, калорийность или другую меру количества живых организмов; 3) пирамида энергии, показывающая величину потока энергии и(или) продукцию на последовательных трофических уровнях (рис. 2).
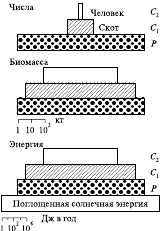
Рис. 2. Трофические пирамиды
Поток энергии через трофический уровень равен общей ассимиляции (А) на этом уровне, а общая ассимиляция, в свою очередь, равна приросту биомассы (продукции – Р) плюс дыхание (R).Экологические пирамиды, построенные по данным численности или биомассам организмов разных трофических уровней, могут быть перевернутыми (обращенными), т. е. с меньшим нижним уровнем по сравнению с некоторыми последующими. Так бывает при сильных различиях в средних размерах организмов продуцентов и консументов, что влечет за собой разную скорость размножения, прироста и образования продукции. Экологическая пирамида энергии, построенная на основе расчета потока энергии или продукции, никогда не будет перевернутой, так как в основу ее построения положены функциональные показатели (прирост, дыхание).
Из-за свойственной большинству видов консументов способности питаться различными видами кормов в биоценозах помимо прямых (вертикальных) пищевых связей возникают боковые (горизонтальные) ответвления, объединяющие потоки вещества и энергии двух и более пищевых цепей, сложным образом переплетающихся и образующих пищевые сети. В пищевой рацион организмов каждого вида обычно входят организмы разных видов, каждый из которых, в свою очередь, может служить пищей организмам других видов. С одной стороны, каждый трофический уровень представлен популяциями разных видов, с другой — организмы многих популяций принадлежат сразу к нескольким трофическим уровням. Таким образом, множественность цепей питания, их разветвленность выступает как приспособление к устойчивому функционированию биоценоза в целом. В результате благодаря сложности пищевых связей выпадение одного вида не нарушает равновесия в экосистеме.
Выделяют два канала поступления и трансформации пищевого материала в экосистемах. Пастбищные цепи – пищевые цепи, начинающиеся с живых фотосинтезирующих организмов (см. приведенный выше пример пищевой цепи пресноводного водоема). Детритные цепи – пищевые цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных.
Пастбищные цепи по масштабам преобладают в водных биоценозах, в которых перерабатывается до 60 % образованной первичной продукции автотрофных организмов (главным образом фитопланктона). В наземных биоценозах цепи разложения преобладают над пастбищными, так как основная масса растительной продукции представлена медленно растущими деревьями и кустарниками, хорошо защищенными от прямого поедания консументами вторичной (пробка) и третичной (корка) покровными тканями, а также наличием большого объема прочной, трудно перерабатываемой древесины. Поэтому около 90 % растительной массы суши перерабатывается в детритных цепях питания.
3. Динамика экосистем. Экологическая сукцессия
Биоценоз зависит от своего биотопа, в равной степени как и биотоп находится под влиянием биоценоза. Поскольку климатические, геологические и биологические факторы подвержены изменениям, развитие биоценозов становится совершенно неизбежным следствием.
Особенно наглядный пример дает ледниковый период на территории средней Европы, когда она представляла собой тундру с карликовыми ивами, дриадами и камнеломками, а вся флора умеренного климата была оттеснена в ее более южные регионы.
Ледниковая фауна включала мамонтов, волосатых носорогов, мускусных овцебыков и мелких грызунов типа леммингов. Таяние и отступление ледников вернуло назад теплолюбивую флору и фауну.
Эрозия, образование осадочных пород, горообразование, вулканическая деятельность, развитие почв могут сильно изменить биотоп и вызвать сдвиги в биоценозе.
Приведенные изменения в биоценозах, обусловленные внешне действующими гео-физико-химическими обстоятельствами, называются аллогенными. В отличие от аллогенных автогенные изменения обусловлены воздействием процессов, протекающих в самих экосистемах. Так, неумеренный выпас коз в Северной Африке демонстрирует замену ранее плодородных биотопов пустынными. Детально описана смена видов животных детритофагов по мере разложения стволов деревьев, трупов животных. Поэтому сообщества постоянно изменяются, сменяя друг друга во времени. Способность к сменам – одно из важнейших свойств живой компоненты экосистем, связанное с тем, что они – открытые системы, находящиеся под влиянием внешних факторов, а также со способностью самого сообщества изменять в результате жизнедеятельности условия своего обитания.
В большинстве случаев трудно разграничить процессы, находящиеся под влиянием внешних (аллогенных) и внутренних (автогенных) факторов. Например, эвтрофикация озер происходит под действием населяющих их сообществ, толчком к изменению которых служит поступление в водоем биогенных веществ извне, с водосбора. Тем не менее степень участия сообщества в преобразовании экосистемы, как правило, устанавливается без особого труда и кроме того, автогенные изменения характеризуются рядом четко обозначенных признаков. Автогенные изменения называют развитием биоценозов или экологической сукцессией, представляющей собой необратимые, т. е. происходящие в определенном направлении изменения биоценоза, проявляющиеся в смене одних видов растений и животных другими. Необратимость и направленность изменений отличает сукцессии от флуктуаций. Направленность изменений при сукцессиях дает основание для предсказания последовательности смен фито- и зооценозов.
Смены, в частности фитоценозов, были давно замечены и описаны. Например, наблюдение за полем, заброшенным на несколько лет без обработки, показывает, что его последовательно заселяют сначала однолетние, затем многолетние травянистые растения, позже кустарниковая и, наконец, древесная растительность. Приоритет в обстоятельной разработке теории этого вопроса принадлежит американским экологам Г. Коулсу и особенно Ф. Клементсу. Ф. Клементс создал систему представлений о сукцессиях, начиная с начальных стадий возникновения биоценоза вплоть до образования устойчивых, самовозобновляющихся и самоподдерживающихся сообществ – климаксов.
Экологическая сукцессия протекает через ряд этапов, при которых происходит замещение одних видов другими под действием модификации среды обитания и создания условий, благоприятных для популяций других видов. Это продолжается до тех пор, пока не будет достигнуто равновесие между биотическими и абиотическими компонентами экосистемы. Последовательность сообществ, сменяющих друг друга, представляет собой серии, названия которых дает преобладающая жизненная форма организмов (например, серия однолетних трав сменяется серией многолетних трав, а затем кустарников и т. д.). В ходе сукцессии лишь немногие виды сохраняются от начальных ее этапов до зрелого (климаксного) состояния экосистемы.
Виды сукцессий. В случае, если развитие экосистемы начинается на пространстве (экотоп), которое никогда не было заселено какими-либо организмами (песчаная дюна, застывший поток лавы, обнажившаяся отмель, скалистая поверхность, породы, обнажившиеся в результате эрозии или отступления ледников), процесс называется первичной сукцессией. Исходное состояние субстрата накладывает отпечаток на прохождение первичных сукцессий, во всяком случае, на их первые стадии.
В ходе первичной сукцессии различают такие последовательно идущие процессы, как образование субстрата (почвы), миграция на него растений и животных, грибов и микроорганизмов, их приживание и агрегация, взаимодействие живой компоненты между собой и с биотопом, изменение сообществом среды, смена биоценозов.
Первые виды в сукцессионном ряду (пионерные) способны заселить малопригодный для жизни субстрат и настолько изменить условия или доступность ресурсов в местообитании, что становится возможным поселение организмов новых видов. Этот процесс известен как облегчение. Он особенно важен при ходе первичной сукцессии в суровых условиях среды. Чаще всего первопоселенцами на каменистых поверхностях либо застывшей магме являются лишайники, так как им, кроме прикрепления, от субстрата нечего не нужно. Медленно растущие и выделяющие лишайниковые кислоты, они постепенно разрушают поверхность субстрата. После отмирания и разложения талломов лишайников образуется перегной, ускоряющий почвообразовательные процессы. Изменение температурного и водного режимов далее дает возможность для поселения мхов, затем травянистых растений, позднее – кустарников и деревьев. При наличии высокой влажности субстратов в сочетании с низкими температурами преимущество среди первопоселенцев переходит к мхам. Ход первичной сукцессии, наблюдаемый в течение 250 лет, с участием мхов как первопоселенцев детально описан при изучении последовательности заселения обнаженной почвы после отступления ледника в районе бухты Глейшер на юго-востоке Аляски (М. Бигон, Дж. Харпер, К. Таунсенд, 1989). Первыми поселенцами были мхи и отдельные сосудистые растения с поверхностной корневой системой, в частности куропаточная трава (Dryas).
Затем появились ивы, сначала стелющиеся, позже кустарниковые виды. Вскоре в сукцессию вовлеклась ольха, и примерно через 50 лет возникли плотные ее заросли высотой до 10 м с отдельными тополями. Постепенно ольха вытеснилась ситхинской елью с примесью тгуса, и в конечном итоге сформировался густой смешанный лес.
Одна из основных движущих сил сукцессии – изменение почвы первыми колонистами. И куропаточная трава, и ольха имеют прокариотические симбионты, фиксирующие атмосферный азот, что приводит к накоплению в почве больших его запасов.
Ольха способствует сильному подкислению почвы, снижая величины рН ее поверхности с 8,0 до 5,0. После этого ситхинская ель, используя накопленный азот, может поселяться и замещать ольху. Постепенное запасание в почве углерода ведет к развитию ее мелкокомковатой структуры, повышению аэрации и влагоемкости субстрата. Таким образом, изменение среды организмами более ранних сукцессионных видов способствует вовлечению других видов, конкурентно вытесняющих своих предшественников.
Экологами было отмечено, что на хуже дренированных склонах при отступлении ледника сукцессия не всегда заканчивалась формированием елово-тгусового леса. Во влажных местах, где уклон отсутствовал или был невелик, бурно развивались сфагновые мхи, накапливающие воду и сильно подкисляющие почву, которая пропитывается влагой и испытывает дефицит кислорода, что приводит к гибели большинства деревьев.
Редко произрастающие сосны (Pinus contorta) –единственные виды деревьев, способные выдержать плохую аэрацию субстрата возникающего болота. Следовательно, завершающая сукцессию растительность явно связана с локальными условиями биотопа.
Длительность периода, необходимого для достижения климаксного состояния, различна. Так, например, в условиях холодного климата Аляски для достижения серии климаксного темнохвойного леса потребовалось 2000 лет. В условиях юго-запада Японии на лавовых отложениях леса достигли климаксного состояния за 700 лет.
Если развитие экосистемы происходит на пространстве, с которого было удалено предыдущее сообщество в результате вырубки или пожара: на заброшенном поле, зарастающем болоте, – то происходит вторичная (восстановительная) сукцессия. Она протекает значительно быстрее, так как сохранена почва с запасами минеральных и органических веществ, необходимых для развития популяций многих видов сообщества.
В качестве примера вторичной сукцессии приведем восстановление елового леса. После вырубки или пожара условия на месте ельника настолько изменяются, что ель не может снова заселить освободившуюся площадь. На открытых местах всходы ели повреждаются поздними весенними заморозками, страдают от перегрева и не могут конкурировать со светолюбивыми растениями. В первые два года на вырубках и гарях буйно развиваются быстрорастущие травянистые растения: кипрей, вейник и др.
Вскоре появляются многочисленные всходы березы, осины, а иногда сосны, т. е. деревьев, семена которых легко разносятся ветром. Деревья вытесняют травянистую растительность и постепенно образуют мелколистный или сосновый лес. Только тогда возникают условия, благоприятные для возобновления ели. Теневыносливые всходы ели успешно конкурируют с подростом светолюбивых лиственных пород. Когда ель достигает верхнего яруса, лиственные деревья постепенно полностью вытесняются, главным образом из-за недостатка света для подроста.
Результаты многочисленных исследований хода процесса сукцессии на залежах свидетельствуют, что типичной сменой доминантных форм растительности является следующая:
однолетние → многолетние → кустарники →раннесукцессионные деревья → позднесукцессионные деревья.
Выявленная последовательность смены жизненных форм растений объясняется рядом их особенностей. Так, семена многих видов однолетних трав сохраняют в почве всхожесть многие годы. Поэтому они, обладая высокой скоростью роста, первыми быстро колонизируют среду залежи, гарантирующую доступ ресурсов в условиях отсутствия конкурентов, появляющихся позже. Смена серии однолетних трав многолетними происходит потому, что быстрое прорастание весной многолетников, имеющих сохранившуюся в зимний период сформированную корневую систему, дает им явное преимущество в росте в весенний период. Многолетники, первыми захватывая неорганические ресурсы почвы и затеняя отставших в росте однолетников, успешно вытесняют последних.
У раннесукцессионных деревьев (береза, осина и др.) расселение семян обычно происходит более эффективно, что повышает вероятность быстрого занятия ими свободного пространства. Они, как правило, скороспелы, т. е. быстро начинают давать семена для занятия новых мест. У позднесукцессионных деревьев семена обычно крупнее и менее эффективно расселяются. Кроме того, раннесукцессионные деревья имеют многоярусную крону, благодаря которой даже в глубине ее освещенность все еще превышает компенсационную точку фотосинтеза. Позднесукцессионные деревья (клен, бук, дуб) можно считать одноярусными породами. У этих деревьев кроны располагаются в один слой, окружающий ствол наподобие шлема. Они успешнее существуют в густом пологе на поздних стадиях сукцессии. Если один и тот же открытый участок одновременно заселяют многоярусные и одноярусные деревья, первые обычно растут быстрее и доминируют до тех пор, пока их не затенят соседи, т. е. пока медленнее растущие одноярусные деревья не перерастут многоярусные.
Один из типов последовательности смен видов при вторичной сукцессии может быть назван деградационной сукцессией, при которой происходит последовательное разложение любой мертвой органики, будь то погибшее животное или растение либо помет. Обычно разные виды сапротрофов появляются и исчезают поочередно, по мере истощения в ходе разложения органического вещества одних ресурсов и появления других веществ. Одновременно изменения в физическом состоянии детрита делают его благоприятным сначала для одних видов, затем для других. Поскольку в этих последовательных сменах участвуют гетеротрофы, сукцессию часто называют гетеротрофной. Она завершается полным потреблением и минерализацией определенного ресурса, осуществляемой редуцентами.
Воспроизведем пример такой сукцессии, описанный в книге М. Бигона, Дж. Харпера, К. Таунсенда (1989). На рис. 3 представлена последовательность колонизации разными видами грибов опавших хвоинок сосны обыкновенной (Pinus sylvestris).
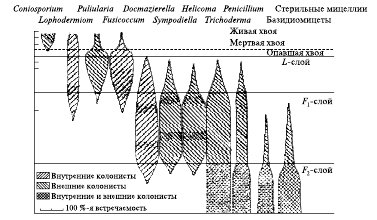
Рис. 3. Пространственно-временные изменения популяций грибов, колонизирующих сосновую хвою в разных слоях лесной подстилки сообщества сосны (Pinus sylvestris) в Англии
Опад непрерывно накапливается под кронами слоями, поскольку земляных червей, зарывающих его в почву, здесь нет. Следовательно, хвоинки на поверхности молодые, а чем глубже они расположены, тем они старше. Сукцессия начинается еще до опадения хвои. Примерно на 50 % живых сосновых хвоинках поселяются организмы рода Сoniosporum. Однако к моменту их старения и отмирания их уже нет, тогда как примерно на 40 % хвоинок поселяются организмы рода Lophodermium, а более 80 % поражены Fusicoccum и Pullularia. Опадающие на землю хвоинки становятся частью так называемого L-слоя, представляющего собой опад из жесткой, еще не слежавшейся светло-бурой хвои. За шесть месяцев пребывания в этом слое большинство хвоинок захватывается Desmazierella. В сукцессионном процессе участвуют и некоторые другие виды, постепенно переваривающие и размягчающие хвою.
Следующая стадия разложения протекает в слое F2 – верхнем подгоризонте ферментации. Серо-черные хвоинки здесь более утрамбованы, их ткани размягчены и непрочны. Внутренняя часть, особенно флоэма, сильно поражена Desmazierella, в то время как внешняя поверхность, окруженная теперь гораздо более влажной атмосферой, заселена Simpodiella и Helicoma. Дальнейшее разрушение внутренних тканей осуществляют почвенные клещи.
Структура хвоинок меняется еще раз. Примерно через два года нахождения в слое F1 они плотно спрессовываются, и участки, ранее пораженные Lophodermium, Fusicoccum и Desmazierella, захватываются различными почвенными животными, главным образом ногохвостками, клещами и олигохетами-энхитреидами. Их деятельность завершает физическое разрушение хвои. В этот период ее частицы медленно заселяются базидиомицетами, способными разрушать целлюлозу и лигнин. Примерно через семь лет пребывания в слое F2 хвоинки делаются структурно неразличимыми, а биологическая активность в очень кислой среде слоя H («гумус») становится крайне низкой.
Большинство организмов перечисленных родов грибов – слабые антагонисты, и время наступления стадий сукцессии зависит, вероятно, главным образом от смены внешних условий и изменений питательных свойств органического вещества хвои.
Эти грибы осуществляют начальное размягчение и переваривание ее тканей, содействуя поселению в хвое и на ней животных-детритофагов. Большая часть пищи, потребляемой последними, вероятно, включает именно грибы, а не растительные остатки.
Приведенный пример разрушения хвои представляет собой конвеерную смену организмов разных таксономических групп, участвующих в деградационной сукцессии.
Климаксное состояние биоценоза. Сукцессия продолжается до тех пор, пока добавление к серии новых видов и исключение предыдущих видов, уже не вызывает изменений среды развивающегося биоценоза. Условия освещения, температуры, влажности и – для первых серий – биогенных веществ почвы быстро изменяются с развитием различных форм роста. На заброшенном поле смена травянистой растительности кустарниками, а затем деревьями сопровождается соответствующими изменениями физической среды. Однако после достижения серии с доминированием деревьев изменение условий среды замедляется и развивается терминальное, стабильное климаксное сообщество. После развития лесной растительности включение в сообщество новых видов деревьев если и изменяет условия влажности почвы и интенсивности освещенности, то в незначительной степени. Сообщество достигает климаксного состояния, так как достигло равновесия со своей абиотической средой.
Однако не все климаксы столь постоянны. Существуют преходящие климаксы.
Примером может быть развитие сообществ растений и животных во временных водоемах, когда сообщество после гибели возобновляется вновь в последующий вегетационный сезон. Каждую весну эти водоемы заселяются либо переселенцами из более крупных постоянных водоемов, либо организмами, развивающимися из спор и покоящихся стадий, оставленных обитателями водоема в предыдущем году до исчезновения местообитания. Здесь сукцессии вновь возобновляются, как только в среде вновь складываются благоприятные для этого условия.
Циклические климаксы формируются тогда, когда вид А развивается после (под сенью) вида Б, вид Б – после вида В, В после вида А. В этом случае создается регулярный цикл доминирования видов в последовательности А-Б-В-А…, причем продолжительность каждой стадии будет определяться продолжительностью жизни соответствующего доминирующего вида. Здесь климакс – динамическое состояние, самовозобновляющееся по составу, даже если это достигается в результате регулярных циклов изменений. Длительное сохранение – ключ к устойчивости. Если цикл сохраняется достаточно долго, то ему присуща такая же стабильность, как и неизменяющемуся стационарному состоянию.
Итак, в климаксном биоценозе достигается равновесие между биотическим сообществом и абиотическими факторами. В результате система приобретает максимальную защищенность от резких изменений среды.
Изучение сукцессии позволяет сформулировать некоторые основные экологические принципы. Во-первых, сукцессия происходит в одном направлении: виды, наделенные способностями к колонизации – быстрым ростом и большой выносливостью к условиям среды в нарушенных или вновь возникших местообитаниях, – сменяются видами с более медленным ростом и высокой способностью к конкуренции. Во-вторых, сукцессионные виды своей структурой и деятельностью изменяют окружающую среду, нередко в ущерб собственной жизни и на благо другим видам. В-третьих, климаксное сообщество не есть нечто единое, а представляет собой в каждом данном месте одну из точек в непрерывном ряду возможных климаксных формаций. На характер и время его достижения оказывают влияние климат, почва, рельеф местности, пожары, деятельность животных.
Биоэнергетическая концепция сукцессий. Представленный выше материал относительно хода экологических сукцессий основан на анализе внешней стороны этого сложного процесса – смене одних видов другими. Каковы же внутренние механизмы экологических сукцессий с точки зрения биоэнергетического подхода? Рассмотрим на конкретном примере образования энергии первичной продукции и закономерностях ее трансформации в экосистемах, находящихся на разных стадиях сукцессионного процесса (табл. 1).
Таблица 1
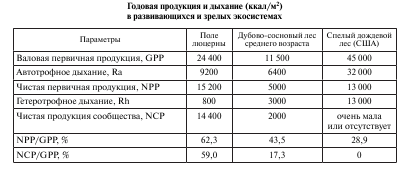
Как видно из таблицы, чистая первичная продукция велика на ранних стадиях сукцессии (поле люцерны) и мала или равна нулю в зрелой системе спелого дождевого леса. Пока P больше R, в системе будет накапливаться органическое вещество и возрастать биомасса (В), в результате чего отношение P/B будет снижаться, а отношения В/Р, В/R или B/E (где E = P + R) – соответственно увеличиваться. Таким образом, мы наблюдаем, что сукцессия связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, затрачиваемой на самоподдержание системы (дыхание), что повышает ее устойчивость.
Анализ большого массива накопленных данных позволил сформулировать биоэнергетическую концепцию хода сукцессии, предложенную известными экологами.
Основные биоэнергетические закономерности сукцессии представлены в табл. 2.
Таблица 2
Биоэнергетическая модель сукцессии экосистем и прогноз ее развития
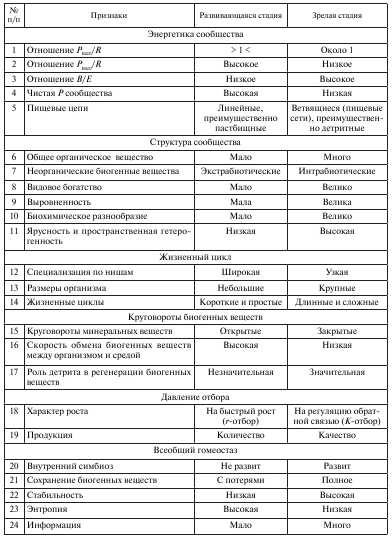
Данные таблицы показывают, что в ходе сукцессии постепенно возрастает видовое разнообразие и выравненность, усложняются связи внутри биоценоза, разветвляются цепи питания и усложняется трофическая сеть, усиливаются симбиотические отношения и регуляторные возможности внутри системы, что уменьшает вероятность слишком сильного размножения отдельных видов, и снижается степень доминирования. Особую значимость имеет отношение энергии, затраченной на поддержание жизнедеятельности (R), к энергии, заключенной в биомассе (В). Это соотношение (R/B) рассматривают как меру термодинамической упорядоченности: чем выше величина этого отношения, тем большей стабильностью обладает биоценоз. Увеличивается КПД всей системы, так как одна и та же порция энергии идет на поддержание большего количества биомассы. Высокая стабильность зрелых биоценозов достигается откачкой энергии на самоподдержание (гомеостаз) – т. е. на дыхание, а не на достижение высокой чистой продукции, поскольку нельзя одновременно сочетать два этих противоположных свойства.
Естественен вопрос: «Стареет ли сама зрелая экосистема, т. е. настанет ли разбалансированность?» В ходе сукцессии и на стадии климакса организмы популяций разных видов, входящие в состав экосистемы, постоянно модифицируют среду, создавая благоприятные условия для организмов других популяций. Это продолжается до тех пор, пока не будет достигнуто равновесие между биотической и абиотической компонентами. Однако не все так однозначно. Накапливаются данные, свидетельствующие о том, что в ряде случаев некоторые виды создают не только условия, благоприятные для других, но и условия, неблагоприятные для самих себя, тем самым ускоряя процесс их замещения другими видами. Так, например, однолетние «сорняки» ранних стадий сукцессии степей и залежных земель нередко продуцируют химические вещества, которые накапливаются в почве и угнетают рост собственных проростков в последующие годы. Процессы, идущие в этом направлении, постепенно приводят к разбалансировке зрелого стабильного биоценоза и постепенной замене его другим.
Изложенная выше концепция с жестко детерминированной сменой сукцессионных серий и однозначным итогом моноклимакса имеет и другие варианты динамики развития биоценозов во времени. При значительном антропогенном вмешательстве либо воздействии природных катаклизмов (интенсивное химическое или радиационное загрязнение, внедрение новых инвазивных видов, длительное подтопление территории, сильные ураганы, смерчи и т. п.) наблюдается нарушение в последовательности и длительности прохождения сукцессионных серий, наличие преходящих поликлимаксов. Новые факторы воздействия на развивающийся биоценоз могут в одном случае поддерживать его организацию, а в другом – быть причиной его разрушения в той или иной степени.
4. ОСНОВНЫЕ ТИПЫ НАЗЕМНЫХ И ВОДНЫХ ЭКОСИСТЕМ
В зависимости от особенностей условий биотопов выделяют наземные и водные экосистемы, которые, в свою очередь, делятся на пресноводные и морские.
Наземные экосистемы. Основными абиотическими факторами, влияющими на размещение по земной поверхности основных наземных экосистем, являются температура и количество осадков. Это формирует особенности климатов в разных районах земного шара, что и определяет наличие климатической зональности в размещении экосистем.
Наземные экосистемы представлены девятью биомами – совокупностями биоты и условий абиотической среды определенной природно-климатической зоны планеты:
тундра – арктическая и альпийская;
тайга – бореальные хвойные леса;
широколиственные леса – листопадный лес умеренной зоны;
степи – злаковники умеренной зоны;
чапараль – районы с дождливой зимой и засушливым летом;
саванна – тропические злаковники;
пустыни – редкая травянистая и кустарниковая растительность;
полувечнозеленый сезонный (листопадный) тропический – в районах с выраженным влажным и сухим сезонами;
джунгли – вечнозеленый тропический дождевой лес.
Водные экосистемы и их основные особенности. Мировой океан является колыбелью жизни. Водные экосистемы самые древние на Земле. Они подразделяются на две основные группы: пресноводные и морские, которые, в свою очередь, делятся на ряд водных экосистем по своим гидрологическим и физическим особенностям.
Морские экосистемы:
открытый океан (пелагическая экосистема);
воды континентального шельфа (прибрежные воды);
районы апвеллинга (зоны подъема глубинных вод, несущие биогенные вещества, создающие высокую продуктивность в эуфотической зоне);
эстуарии (прибрежные бухты, проливы, эстуарии, лиманы, соленые марши);
глубоководные рифтовые зоны.
Пресноводные экосистемы:
лентические (стоячие воды) – озера, пруды, водохранилища;
лотические (текущие воды) – реки, ручьи, родники;
заболоченные угодья – болота, марши (приморские луга).
Озера и водохранилища относятся к лентическим экосистемам. В них в литоральной (прибрежной) зоне преобладают две группы растений: укрепившиеся на дне (камыши, рогозы, кувшинки, прикрепленные водоросли и др.) и плавающие (ряска, некоторые водоросли). В этой зоне встречаются моллюски, мшанки, личинки насекомых. Рыбы большую часть жизни проводят в литоральной зоне, животные (в том числе и вторичноводные животные) дышат кислородом атмосферного воздуха (лягушки, прудовики, некоторые водные паукообразные). Зоопланктон представлен ракообразными (дафниями, циклопами и др.), имеющими большое значение в питании личинок и молоди рыб.
В лимнической зоне продуценты представлены планктонными фотоавтотрофными протистами и цианобактериями, а зоопланктон – растительноядными (фильтраторами) и хищными ракообразными, а также коловратками. Нектон лимнической зоны – только рыбы.
Профундальная (бессветовая, донная) область представлена бентосными формами – личинками насекомых, моллюсками, малощетинковыми кольчатыми червями, сапротрофными бактериями и грибами.
К лотическим экосистемам относятся реки. Они отличаются от стоячих водоемов наличием течения, более активным обменом газами с атмосферой, высоким содержанием кислорода и равномерным его распределением по вертикальному профилю реки, а также преобладанием детритных цепей питания. В реках выделяют лотические сообщества перекатов и плесов. На перекатах встречаются организмы, способные прикрепиться к субстрату (например, ручейники, нитчатые водоросли), или хорошие пловцы (форель). На участках плеса сообщества напоминают озерные. В больших реках наблюдается продольная зональность: в верховьях – сообщества перекатов, в низовье и дельте – сообщества плесов, а между ними могут возникать и те, и другие.
| <== предыдущая лекция | | | следующая лекция ==> |
| Виды электрических кабелей и проводов | | | ОРГАНИЗМ КАК ОБЪЕКТ ЭКОЛОГИИ |
Дата добавления: 2022-04-12; просмотров: 141;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
