Филогенез скелета позвоночных.
Скелет позвоночных образуется из мезодермы и состоит из 3 отделов: скелета головы (череп), осевого скелета туловища (хорда, позвоночник и ребра), скелета конечностей и их поясов.
Основные направления эволюции осевого скелета:
1. Замена хорды позвоночником, хрящевой ткани – костной.
2. Дифференцировка позвоночника на отделы (от двух до пяти).
3. Увеличение числа позвонков в отделах.
4. Формирование грудной клетки.
Круглоротые и низшие рыбы сохраняют хорду в течение всей жизни, но у них уже появляются зачатки позвонков (парные хрящевые образования, расположенные над и под хордой): верхние дуги у круглоротых, и нижние – у рыб.
У костных рыб развиваются тела позвонков, появляются остистые и поперечные отростки, формируется канал спинного мозга. Позвоночник состоит из 2 отделов: туловищного и хвостового. В туловищном отделе есть ребра, которые свободно заканчиваются на брюшной стороне тела.
У амфибий появляются 2 новых отдела: шейный и крестцовый, каждый из них содержит по одному позвонку. Имеется хрящевая грудина. Ребра у хвостатых амфибий незначительной длины и никогда не доходят до грудины, у бесхвостых амфибий ребра отсутствуют.
В позвоночнике рептилий выделяют шейный отдел, который содержит 8-10 позвонков, грудной, поясничный (в этих отделах – 22 позвонка), крестцовый – 2 и хвостовой, в котором может быть несколько десятков позвонков. Первые два шейных позвонка имеют особое строение, в результате чего обеспечивается большая подвижность головы. Три последних шейных позвонка имеют по паре ребер. Первые пять пар ребер пояснично-грудного отдела присоединяются к хрящевой грудине, образуя грудную клетку.
У млекопитающих позвоночник состоит из 5 отделов. Шейный отдел имеет 7 позвонков, грудной – от 9 до 24, поясничный – от 2 до 9, крестцовый – 4-10 и более, в хвостовом отделе – очень большие вариации. Происходит редукция ребер в шейном и поясничном отделах. Грудина костная. 10 пар ребер доходят до грудины, формируя грудную клетку.
Онтофилогенетически обусловленные аномалии скелета: дополнительные ребра у седьмого шейного или у первого поясничного позвонка, расщепление задней дуги позвонков, несрастание остистых отростков позвонков (Spinabifida), увеличение числа крестцовых позвонков, наличие хвоста и др.
Череп позвоночных развивается как продолжение осевого скелета (мозговой отдел) и как опора для дыхательной и передней части пищеварительной систем (висцеральный отдел).
Основные направления эволюции черепа:
1. Объединение висцерального (лицевого) отдела с мозговым, увеличение объема мозгового отдела.
2. Уменьшение числа костей черепа за счет их слияния.
3. Замена хрящевого черепа костным.
4. Подвижное соединение черепа с позвоночником.
Происхождение осевого черепа связано с метамерией (сегментацией) головы. Его закладка происходит из двух основных отделов: хордального – по бокам от хорды, который сохраняет расчленение на сегменты (парахордалии), прехордального – впереди хорды (трабекулы).
Трабекулы ипарахордалии разрастаются и сливаются вместе, образуя черепную коробку снизу и с боков. К ней прирастают обонятельные и слуховые капсулы. Боковые стенки заполняют глазничные хрящи. Осевой и висцеральный череп развиваются по-разному и на ранних этапах фило- и онтогенеза не связаны между собой. Мозговой череп проходит три стадии развития: перепончатую, хрящевую и костную.
У круглоротых крыша мозгового черепа соединительнотканная (перепончатая), а основание образовано хрящевой тканью. Висцеральный череп представлен скелетом предротовой воронки и жаберным, состоящим у миног из ряда семи хрящей.
У низших рыб осевой череп хрящевой (рисунок 8). Появляется затылочный отдел. Висцеральный череп состоит из 5-6 метамерно расположенных хрящевых дуг, которые охватывают передний отдел пищеварительной трубки. Первая дуга, самая крупная, называется челюстная дуга. Она состоит из верхнего хряща – небноквадратного, который образует первичную верхнюю челюсть. Нижний хрящ – меккелев хрящ, образует первичную нижнюю челюсть. Вторая жаберная дуга – подъязычная (гиоидная), состоит из двух верхних гиомандибулярных хрящей и двух нижних – гиоидов. Гиомандибулярный хрящ с каждой стороны срастается с основанием мозгового черепа, гиоид соединяется с меккелевым хрящем. Таким образом челюстная дуга соединяется с мозговым черепом и такой тип соединения висцерального и мозгового черепа называется гиостильный.
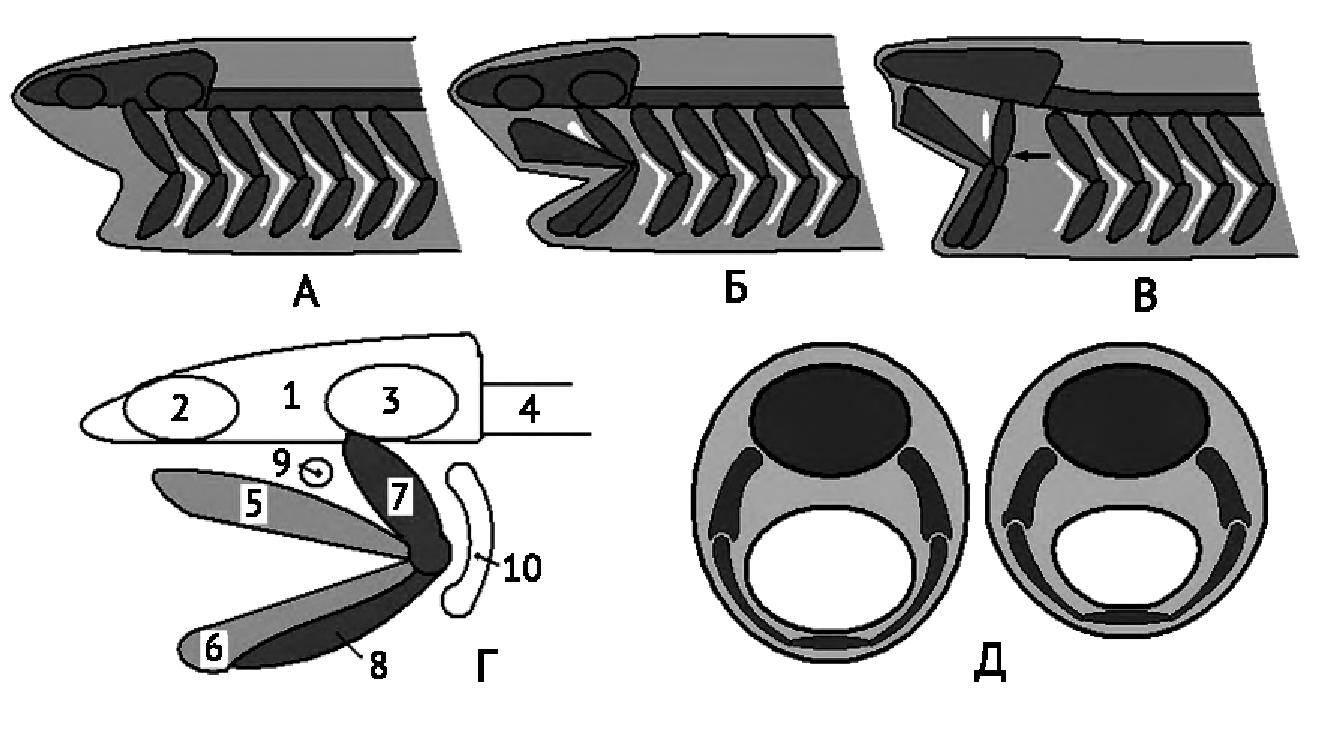
Рисунок 8. Челюсти (по Ромер, Парсонс, 1992). А-В – видоизменение первых двух пар жаберных дуг в челюсти рыб; Г – скелет головы акулы: 1 — череп, 2 — обонятельная капсула, 3 — слуховая капсула, 4 — позвоночник, 5 — нёбно- квадратный хрящ (верхняя челюсть), 6 — меккелев хрящ, 7 — гиомандибуляре, 8 — гиоид, 9 — брызгальце (первая недоразвитая жаберная щель), 10 — первая полная жаберная щель: Д – поперечный срез акулы в области головы.
У костных рыб развивается вторичный костный череп. Он частично состоит из костей, которые развиваются из хрящей первичного черепа, а также из покровных костей, которые прилегают к первичному черепу. Крыша мозгового черепа состоит из парных лобных, теменных и носовых костей. В затылочной области имеются затылочные кости. В висцеральном черепе из покровных костей развиваются вторичные челюсти. Роль верхней челюсти переходит к покровным костям, которые развиваются в верхней губе, нижней челюсти, а также к покровным костям, развивающимся в нижней губе. На других висцеральных дугах покровные кости не развиваются. Тип соединения мозгового и висцерального черепа гиостильный. Череп у всех рыб неподвижно соединяется с позвоночником.
Череп наземных позвоночных изменяется, главным образом, в связи с утратой жаберного дыхания. У земноводных в мозговом черепе сохраняется еще много хрящей, он становится легче черепа рыб. Характерным для всех наземных позвоночных является подвижное соединение черепа с позвоночником. Наибольшие изменения происходят в висцеральном черепе. У амфибий функционируют вторичные челюсти. Первая, челюстная дуга, частично редуцируется. Небноквадратный хрящ первой челюстной дуги срастается с основанием мозгового черепа – такой тип соединения называется аутостильный. В связи с этим гиомандибулярный хрящ гиоидной дуги теряет роль подвеска челюстной дуги. Он преобразуется в слуховую косточку (столбик), расположенную в слуховой капсуле. Нижний хрящ первой жаберной дуги – меккелев хрящ – частично редуцируется, а оставшаяся часть его окружена покровными костями. Гиоид (нижний хрящ второй дуги) преобразуется в передние рожки подъязычной кости. Остальные висцеральные дуги (всего у амфибий их 6) сохраняются в виде подъязычной кости и в виде гортанных хрящей.
У рептилий череп взрослого животного окостеневает. Имеется большое число покровных костей. Соединения висцерального и мозгового черепа происходит за счет квадратной кости (окостеневшей задней части редуцированного небноквадратного хряща). Череп аутостильный. Челюсти вторичные. Изменени других частей висцеральных дуг такие же, как у амфибий. У рептилий образуется вторичное твердое небо и скуловые дуги.
У млекопитающих наблюдается уменьшение числа костей в результате их срастания и увеличение объема мозгового черепа. Крыша черепа образована лобными и теменными костями, височная область прикрыта скуловой дугой. Вторичные верхние челюсти образуют переднюю нижнюю часть черепа. Нижняя челюсть состоит из одной кости и ее отросток образует сустав с помощью которого она соединяется с мозговым черепом.
Рудименты небноквадратного и меккелева хряща преобразуются соответственно в слуховые косточки – наковальню и молоточек. Верхний отдел подъязычной дуги образует стремечко, нижний отдел – подъязычный аппарат. Части 2-ой и 3-я жаберные дуги образуют щитовидный хрящ гортани, 4-ая и 5-ая дуги преобразуются в остальные хрящи гортани. У высших млекопитающих значительно увеличивается объем мозгового черепа. У человека размеры лицевого черепа значительно уменьшаются по сравнению с мозговым отделом, черепная коробка округлая и гладкая. Формируется скуловая дуга (синапсидный тип черепа).
Онтофилогенетически обусловленные пороки черепа: увеличение числа костных элементов (каждая кость может состоять из большого числа костей), несращение твердого неба – "волчья пасть", лобный шов, верхняя часть затылочной чешуи может быть отделена от остальной части поперечным швом; в верхней челюсти встречается непарная резцовая кость характерная для других млекопитающих, одна слуховая косточка, отсутствие подбородочного выступа и др.
Основные направления эволюции скелета поясов и свободной конечности:
1. От кожных (метаплевральных) складок ланцетника к парным плавникам рыб.
2. От многолучевого плавника рыб к пятипалой конечности.
3. Увеличение подвижности соединения конечностей с поясами.
4. Уменьшение числа костей свободной конечности и укрупнение их путем слияния.
Основой образования конечностей позвоночных являются кожные складки по бокам туловища (метаплевральные), которые имеются у ланцетника и личинок рыб.
В связи с изменением функции метаплевральные складки изменили свое строение. У рыб в них появились мышцы и скелет, в виде метамерного ряда хрящевых лучей, образующих внутренний скелет плавников. У высших рыб плавниковые лучи костные. Первичный передний пояс представляет собой дугу (в основном, костную), которая охватывает тело с боков и с брюшной стороны. Пояс лежит поверхностно, покрыт несколькими костями, гомологичными лопатке и коракоиду высших позвоночных. Он служит только для связи плавников с вторичным поясом. Вторичный пояс состоит из крупной парной кости, которая на спинной стороне прикрепляется к крыше черепа, а на брюшной – соединяется между собой. Задний пояс рыб развит слабо. Он представлен небольшой парной пластинкой. У кистеперых рыб плавники стали служить опорой при передвижении по грунту и в них произошли изменения, которые подготовили их к преобразованию в пятипалую конечность наземных позвоночных (рисунок 9). Уменьшилось число костных элементов, они стали крупнее: проксимальный отдел составляет одна кость, средний – две кости, дистальный – радиально расположенные лучи (7 – 12). Сочленение скелета свободной конечности с поясами конечностей стало подвижным, что позволило кистеперым рыбам использовать плавники как опору для тела при передвижении по грунту.
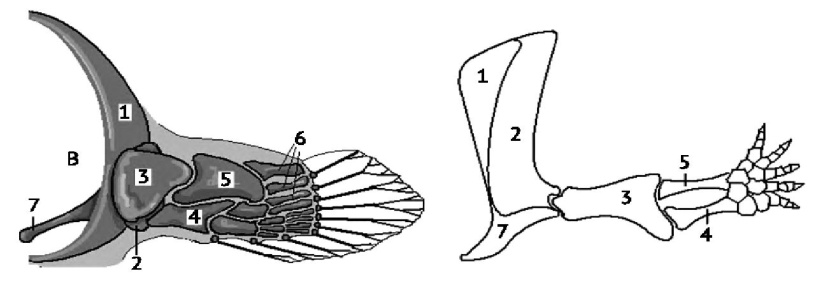
Рисунок 9. Грудной плавник кистеперой рыбы и передняя лапа древней амфибии (по Кэрролл, 1992). 1 — клейтрум, 2 — лопатка, 3 — базалия, соответствующая плечевой кости, 4 — базалия, соответствующая локтевой кости, 5 — базалия, соответствующая лучевой кости, 6 — радиалии, 7 — ключица.
Следующий этап эволюции – замена прочного соединения элементов скелета подвижными сочленениями, уменьшение у высших позвоночных числа рядов в запястье и числа костей в ряду, значительное удлинение проксимального (плеча, предплечья) и дистальных отделов (пальцев), а также укорочение костей среднего отдела.
Конечность наземных позвоночных представляет собой сложный рычаг, который служит для перемещения животного на суше. Пояса конечностей (лопатки, вороньи, ключицы) имеют вид дуги, которая охватывает тело с боков и снизу (рисунок 10). Для присоединения свободной конечности на лопатке имеется углубление, а сами пояса становятся более широкими, что связано со значительным развитием мускулатуры конечностей. У наземных позвоночных тазовый пояс состоит из 3-х парных костей: подвздошных, седалищных и лобковых (рисунок 11).Седалищные кости соединяются с крестцом. Все три кости образуют вертлужную впадину. Хорошо развит спинной отдел поясов, что способствует их более прочному укреплению.
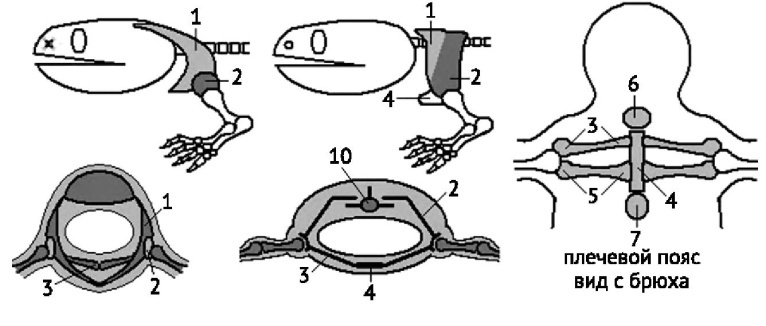
Рисунок 10. Сравнение поясов передних конечностей кистепёрых рыб (слева) и амфибий (справа) (по Квашенко, 2014). 1 — клейтрум, 2 — лопатка, 3 — ключица, 4 — грудина, 5 — коракоид, 6 — предгрудина, 7 — загрудина.
У человека встречаются онтофилогенетически обусловленные аномалии скелета конечностей: плоскостопие, добавочные кости запястья, предплюсны, добавочные пальцы кисти или стопы (полидактилия) и др.
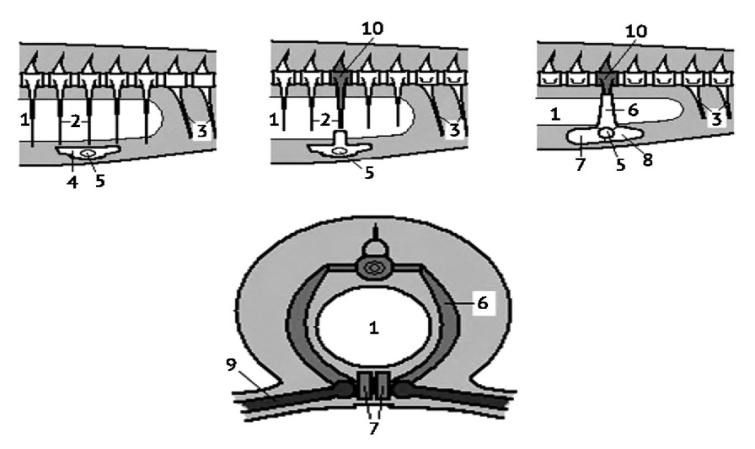
Рисунок 11. Развитие тазового пояса наземных позвоночных в связи с редукцией рёбер (по Квашенко, 2014). 1 — целом, 2 — ребра, 3 — брюшные остистые отростки, 4 — тазовая пластинка рыб, 5 — ямка тазобедренного сустава, 6 — подвздошная кость, 7 — лобковая кость, 8 — седалищная кость, 9 — бедренная кость, 10 — крестцовый позвонок.
Дата добавления: 2016-05-28; просмотров: 16390;
Поиск по сайту
Узнать еще
- анатомия костей скелета. соединениякостей
- Аномалии развития скелета и структурных дефектов у собак
- Биологических структур в филогенезе
- Гисто- и орагногенез. Процесс нейруляции. Осевые органы и их формирование. Дифференцировка мезодермы. Производные органы зародышей позвоночных.
- Значение опорно-двигательного аппарата. Химический состав костей. Строение скелета головы, туловища, конечностей. Виды соединений костей. Строение сустава.
- Мускулатура осевого скелета
- Мышцы периферического скелета
- Общее представление о механизмах развития скелета
Публикации по технике и механике
Публикации по биологии
Публикации по информатике
Публикации по строительству
Публикации по физике
Публикации по химии
Публикации по электронике

Публикации по искусству

Публикации по истории