Половые и соматические клетки. Их сходство и различия
Все современные многоклеточные организмы состоят из генеративной (половые клетки) и соматической (из которой развиваются все остальные органы) частей. Подобное разделение является важнейшим эволюционным событием, которое и обусловило переход от одно- к многоклеточности и сделало возможным сам процесс онтогенеза, сводящийся главным образом к прогрессивному усложнению и специализации соматической части организма.
Основные отличия половых клеток от соматических
1.Сперматозоиды и яйцеклетки имеют гаплоидный набор хромосом, а не диплоидный, как это свойственно соматическим клеткам.
2.Для половых клеток характерно сложное, стадийное развитие; при этом имеет место особый способ деления – мейоз.
3.Половые клетки тотипотентны, т. е. они сохраняют способность формировать любые (все) органы и ткани организма. Если из соматической клетки может образоваться лишь такая же дочерняя клетка, то из половых клеток формируется целый новый организм.
4.У половых клеток по сравнению с соматическими резко изменено ядерно-плазменное отношение: у яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша, а у сперматозоидов благодаря малому количеству цитоплазмы ядерно-цитоплазматическое отношение высокое. Это находится в соответствии с главной функциональной задачей мужской гаметы — транспортировкой наследственного материала к яйцеклетке. Впоследствии при развитии зародыша ядерно-плазменное отношение делящихся клеток восстанавливается до свойственного соматическим клеткам. Это происходит у разных животных в неодинаковое время, но большей частью к 5-7 делению яйца.
5.Различный уровень метаболизма: яйцевая клетка в отношении обмена веществ находится в состоянии депрессии, а у сперматозоидов столь малое количество цитоплазмы и питательных веществ, что нормальный метаболизм и вовсе исключается. В половых железах или половых протоках самца сперматозоиды находятся в неподвижном анабиотическом состоянии. Оказавшись вне мужской половой системы, они живут очень короткое время. Однако есть исключения и из этого правила. Например, у летучей мыши спаривание происходит осенью, но оплодотворения при этом не происходит. Вскоре животные впадают в спячку, при этом сперматозоиды сохраняются в течение зимнего периода в половых путях самок, а лишь весной происходит оплодотворение;
6.Яйцеклетки и сперматозоиды представляют собой высокоорганизованные клетки, обладающие многими специальными, выработанными в процессе эволюции приспособлениями для выполнения специфических функций (жгутик, оболочки яйцеклеток); – ♂ имеет акросому (для проникновения через оболочки ♀ ) и мощный двигательный аппарат – хвостик;
– ♀ яйцеклетка имеет желток (запас питательных веществ и строительных материалов) и оболочки (I, II, а у некоторых видов и III).
7.Сперматозоиды неспособны развиваться и доходить до итогового этапа в жизни клетки — митоза. Яйцеклетки также не могут делиться без влияния особых факторов: если не произойдет оплодотворение или если они не будут активированы к развитию партеногенетическими агентами.
Половые клетки в своем развитии претерпевают ряд сложных преобразований.
При образовании гамет происходит деление клетки, называемое мейозом. Исходная клетка имеет диплоидный набор хромосом, которые затем удваиваются. Но, если при митозе в каждой хромосоме хроматиды просто расходятся, то при мейозе хромосома (состоящая из двух хроматид) тесно переплетается своими частями с другой, гомологичной ей хромосомой (также состоящей из двух хроматид), и происходит кроссинговер - обмен гомологичными участками хромосом. Затем уже новые хромосомы с перемешанными «мамиными» и «папиными» генами расходятся и образуются клетки с диплоидным набором хромосом, но состав этих хромосом уже отличается от исходного, в них произошла рекомбинация. Завершается первое деление мейоза, и второе деление мейоза происходит без синтеза ДНК, поэтому при этом делении количество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидных клетки.
Строение спермия
♂ - самая маленькая клетка в организме, у различных животных сильно отличаються в строении. Пребладающей формо являются бичевидные.♂ состоит из
ü Головки, состоящей из секреторного пузырка – акросомы (содержащий
гидролитические ферменты, и позволяющий спермию проникнуть через
наружные яйцевые оболочки) и ядра (содеожит мужской наследственный
материал в виде протного хроматина). Головка ♂-а окружена очень
тонким слоем цитоплазмы. Когда головка спермия приходит в контакт с
♀-ой происходит акросомальная реакция - освобождение содержимого
акросомы путем экзоцитоза.
ü Шейки, содержащую проксимальнуюи дистальную центриоли,
расположенные перпендикулярно друг к другу;
ü Средней части, содержащая пучок фибрилий (2-х центральных и 9-ти пар
периферических), митохондрий, спирально расположенных вокруг осевой
нити. Эта часть обеспечивает метаболическую и энергетическую
активность ♂;
ü Хвоста, содержит осевую нить, окруженную ебольшым количеством
цитоплазмы и клеточной (ундулирующей) мембраной. Передвижение
осуществляется путем сгибально-разгибательных, ударных и
волнообразных движений. ♂ многих животных лишены хвота.
На ♂ имеется для выбора направления движения хеморецепторы,
сходные с обонятельными клетками
Каждый спермий содержит: гаплоидное ядро; двигательную систему, обеспечивающую перемещение ядра, и пузырек, заполненный ферментами, необходимыми для проникновения ядра в яйцо (рис. 1).
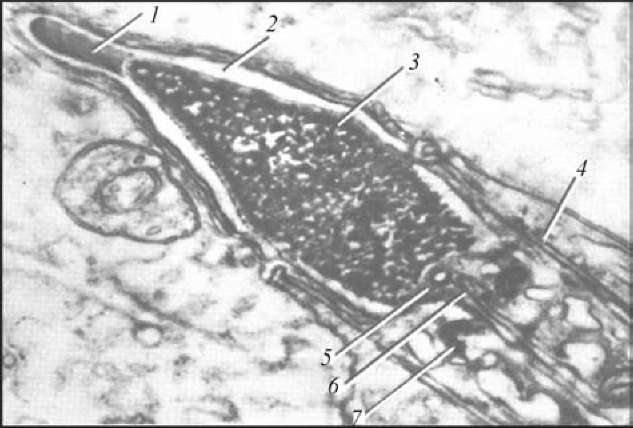 Рис. 1. Строение поздней сперматиды (по Алмазову, Сутулову, 1978):
1 — акросома; 2 — головной колпачок; 3 — ядро; 4 — цитоплазма; 5 — проксимальная центриоль; 6—осевые нити; 7 — дистальная центриоль
Рис. 1. Строение поздней сперматиды (по Алмазову, Сутулову, 1978):
1 — акросома; 2 — головной колпачок; 3 — ядро; 4 — цитоплазма; 5 — проксимальная центриоль; 6—осевые нити; 7 — дистальная центриоль
|
Большая часть цитоплазмы спермия элиминируется при его созревании. Сохраняются только некоторые органеллы, видоизмененные для выполнения своей функции. В период созревания спермия его гаплоидное ядро приобретает обтекаемую форму, а ДНК уплотняется. Впереди от такого конденсированного гаплоидного ядра лежит акросомный пузырек, образующийся из аппарата Гольджи и содержащий ферменты, которые переваривают белки и полисахариды. Запас ферментов в акросомном пузырьке служит для проникновения спермия через наружные покровы яйца. У морских ежей между ядром и акросомным пузырьком находится область, содержащая глобулярный актин. Он используется для образования пальцеобразного выроста. У таких видов молекулы на поверхности акросомного выроста участвуют в узнавании спермием и яйцом друг друга. Акросома и ядро образуют вместе головку спермия.
Акросома, производное аппарата Гольджи, имеет свою мембрану, в которой выделяют следующие части: наружную, промежуточную, внутреннюю (прилежащую к ядру), в последней выделяют инвагинационные трубочки, их 15. Внутри акросомы находится акросомальная гранула, она не имеет своей мембраны. Внутри акросомы есть ферменты: гиалуронидаза и трипсин. Они воздействуют на оболочку яйцеклетки: гиалуронидаза растворяет блестящую оболочку яйцеклетки, трипсин нарушает целостность фолликулярной оболочки.
У большинства видов спермии способны передвигаться на большие расстояния благодаря биению своих жгутиков (рис. 2).
Главная двигательная основа жгутика — аксонема. Она берет начало от дистальной центриоли, которая находится в шейке. Осевая нить проходит через весь вставочный отдел и через весь хвостик. Во вставочном отделе вокруг аксонемы находится спиральная структура, которая образуется 12—15 витками митохондрий. Стержень аксонемы состоит из двух центральных одиночных микротрубочек, окруженных кольцом из девяти двойных микротрубочек (дуплетов). При этом только одна микротрубочка каждого дуплета имеет законченное строение и содержит 13 протофиламентов, тогда как вторая состоит из 11 протофиламентов димерного белка тубулина. С микротрубочками связан белок динеин. С его помощью гидролизуются молекулы АТФ и преобразуется выделившаяся при этом химическая
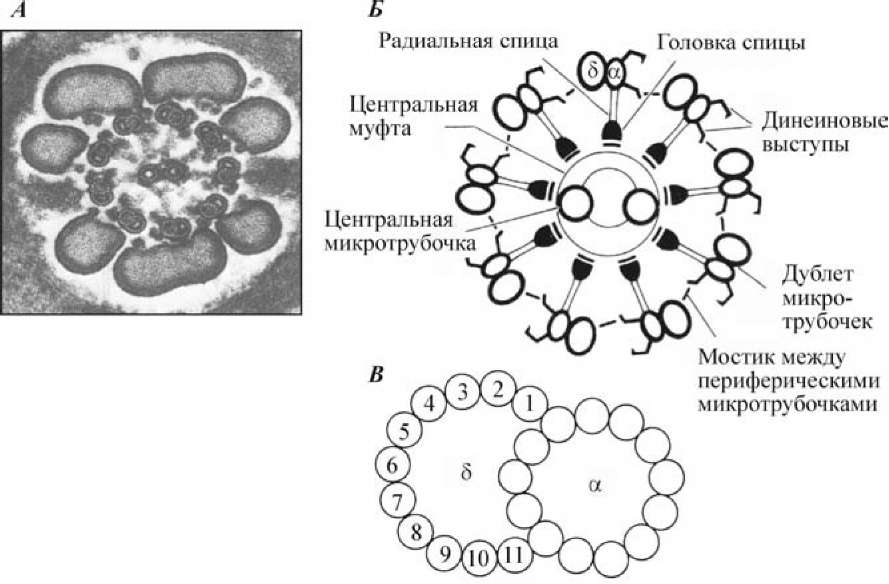 Рис. 2. Строение хвостика сперматозоида (по Гилберту, 1993): А — поперечный срез жгутика млекопитающего. Видны центрально расположенная аксонема и наружные волокна; Б — схема строения аксонемы. Комплекс микротрубочек «9+2», радиальные спицы, динеиновые выступы и др.; В — расположение микрофиламентов тубулина в дуплете микротрубочек. Часть дуплета а является нормальной микротрубочкой, содержащей 13 микрофиламентов. Часть 8 содержит 11 микрофиламентов
Рис. 2. Строение хвостика сперматозоида (по Гилберту, 1993): А — поперечный срез жгутика млекопитающего. Видны центрально расположенная аксонема и наружные волокна; Б — схема строения аксонемы. Комплекс микротрубочек «9+2», радиальные спицы, динеиновые выступы и др.; В — расположение микрофиламентов тубулина в дуплете микротрубочек. Часть дуплета а является нормальной микротрубочкой, содержащей 13 микрофиламентов. Часть 8 содержит 11 микрофиламентов
|
энергия в механическую, за счет которой осуществляется движение спермиев. Мужчины с генетическим синдромом отсутствия динеина во всех клетках, обладающих ресничками и жгутиками, характеризуются следующим (триада Картедженера): они стерильны (из-за неподвижности спермиев), подвержены респираторным инфекциям (из-за неподвижности ресничек мерцательного эпителия, выстилающего дыхательные пути), у них в 50 % случаев сердце располагается с правой стороны.
На поперечном срезе через аксонему видны фибриллы — в центре 2 центральные фибриллы, по периферии 9 пар периферических субфибрилл, всего их 20, они связаны между собой структурами, которые называются спицами. Центральные субфибриллы выполняют функцию проведения, периферические — сокращения. Так как во вставочном отделе имеются митохондрии, сперматозоиды способны к самостоятельным движениям. Скорость движения составляет 2-5 мм/мин. Движение сперматозоидов против тока секрета носит название реотаксиса. Направление движения: вперед-вверх или вперед-вниз, вращение вокруг собственной оси. Размеры сперматозоидов составляют: морской свинки — 100 мкм, быка — 65 мкм, воробья — 200 мкм, крокодила — 20 мкм, человека — 60 мкм. Для обеспечения оплодотворения необходимо, чтобы в 1 мл спермы человека содержалось около 60 млн сперматозоидов.
Оогенез
Яйцевые клетки образуются в женской половой железе — яичнике (ovarium), расположенной в области малого таза, длиной 2,5-5,5 см, шириной 1,5-3,0 см, толщиной до 2 см, весом 5-8 г. Они проходят долгий путь развития, который начинается в эмбриональном и продолжается в репродуктивном периоде онтогенеза особей женского пола (рис.).
Первичные половые клетки возникают на ранних этапах эмбриогенеза из энтодермальных клеток вегетативного полюса, как, например, у бесхвостых амфибий, или из энтодермальных клеток желточного мешка, как у всех амниот — пресмыкающихся, птиц и млекопитающих. ППК очень рано отличаются от других клеток благодаря крупным размерам и прозрачной цитоплазме. Половые железы в этот момент только начинают формироваться. Экспериментально было показано, что первичные половые клетки мигрируют из места возникновения в развивающиеся гонады и заселяют их. У млекопитающих они перемещаются по дорсальной брыжейке, будучи способными в этот период к амебоидному движению. У птиц миграция происходит пассивно по кровяному руслу. У высших позвоночных не обнаружено веществ, стимулирующих миграцию первичных половых клеток к гонадам. Существует мнение, что первичные половые клетки, оказавшись в каких-либо других участках зародыша, как правило, гибнут, но иногда могут перерождаться в опухоли.
Попав в гонады, первичные половые клетки начинают пролиферировать. Они делятся митозом и называются оогониями. Наступает стадия размножения. У большинства низших позвоночных оогонии сохраняют способность к делению на протяжении всего репродуктивного периода, например, рыбы за один нерест выделяют тысячи яиц, земноводные — сотни (животные с наружным оплодотворением).
Виды, для которых характерно внутреннее оплодотворение, продуцируют половые клетки более экономно. У млекопитающих размножение овогониев протекает только в эмбриональном периоде и к концу внутриутробного развития прекращается. Так, у человека максимальное количество оогониев (6-7 млн) наблюдается у пятимесячного плода. Далее следует массовая дегенерация половых клеток, количество которых у новорожденной девочки составляет около 1 млн, а к семи годам сокращается до 300 тысяч.
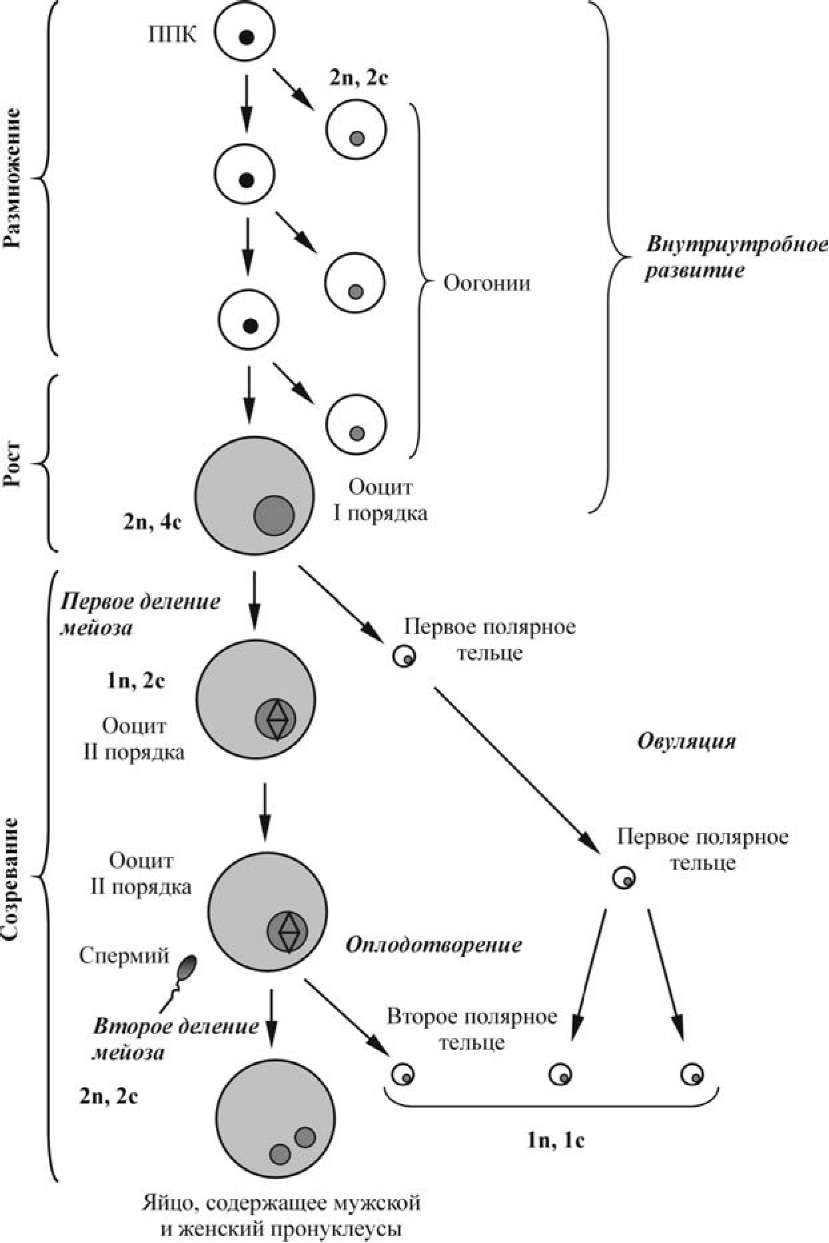 Рис. Схема процесса оогенеза человека
Рис. Схема процесса оогенеза человека
|
Женская половая клетка, прекратившая размножение, называется ооцит I порядка. Начинается свойственный только этой клетке период роста. Он связан с поступлением в яйцеклетку питательных веществ извне и с синтезом их в самой яйцеклетке. Масса и объем яйцеклетки увеличиваются в огромное количество раз (у насекомых — в 90 000 раз, у млекопитающих — более чем в 40 раз).
Рост ооцитов принято разделять на два периода:
• малого, или цитоплазматического, роста (превителлогенез): происходит относительно небольшое пропорциональное увеличение массы ядра и цитоплазмы;
• большого, или трофоплазматического, роста (вителлогенез): резко интенсифицируется рост цитоплазматических компонентов, в ооците откладывается желток.
Весь период превителлогенеза проходит на фоне подготовки ооцита I порядка к последующим делениям созревания (мейоза). Подготовка к первому делению созревания начинается с того, что ооцит вступает в S-период редукционного деления (фаза удвоения ДНК). Затем следует профаза первого деления мейоза, которая продолжается у ооцитов млекопитающих несколько дней.
По достижении фазы диплотены, когда гомологичные хромосомы уже прошли конъюгацию и начали расходиться к противоположным полюсам ядра, наступает стадия диакинеза. На ней дальнейшее течение мейоза сильно замедляется. Остановка мейоза продолжается до достижения особью по- ловозрелости, т. е. длится много лет, применительно к некоторым млекопитающим и человеку. В период диакинеза ядерный материал ооцита не остается инертным: у большинства яйцеклеток он выполняет роль матрицы для синтеза всех видов РНК — информационных, транспортных, матричных и рибосомальных. Все эти типы РНК синтезируются впрок и используются уже оплодотворенной яйцеклеткой. Синтез рРНК связан с уникальным процессом амплификации генов (т. е. временного увеличения числа генов, кодирующих данный вид РНК). Амплификация осуществляется путем избирательного копирования рибосомальных генов, расположенных вдоль нити ДНК. Отделившиеся копии обособляются морфологически в виде ядрышек, которых может насчитываться несколько тысяч.
После созревания ооцита ядрышки выходят в его цитоплазму и там ли- зируются. Синтез рРНК протекает в течение 3-6 месяцев. Низкомолекулярные рРНК и тРНК синтезируются без амплификации — их быстрое накопление обусловлено тем, что кодирующие их гены многократно повторены. Усиливающаяся синтетическая активность нуклеиновых кислот приводит к образованию хромосом типа ламповых щеток, что связано с наличием деспирализованных участков ДНК, на которых идет синтез мРНК. В зрелой яйцеклетке насчитывается до 25-50 тыс. различных типов мРНК.
В период вителлогенеза в ооците I порядка образуется желток, а также жиры и гликоген. Желток — это высокофосфорилированный кристаллический белок. Его количество в клетке строго детерминировано генетически и не зависит от условий питания самки. Вителлогенез может осуществляться за счет синтеза желтка внутри ооцита (эндогенный желток), либо желток синтезируется вне яичника (экзогенный желток). Синтез эндогенного желтка происходит в эндоплазматическом ретикулуме из концевых цистерн аппарата Гольджи. Накопление желтка может идти и в митохондриях, которые при этом перерождаются в желточные гранулы. Большинству видов животных присуще образование экзогенного желтка. Он строится на основе белка, предшественника вителлогенина, поступающего в ооцит извне.
У позвоночных вителлогенин синтезируется в печени матери, транспортируется к содержащему ооцит фолликулу по кровеносным сосудам и поглощается ооцитом путем пиноцитоза. В дальнейшем, при формировании гранул желтка, он распадается на липовителлин и фосфовитин, входящие в состав экзогенного желтка. Синтез вителлогенина клетками печени находится под гормональным контролем. Выделяемый гипоталамусом лю- либерин стимулирует продукцию гонадотропных гормонов гипофиза (ФСГ, ЛГ) в кровь. Под их воздействием клетки фолликула синтезируют в кровоток эстроген. Последний индуцирует, а впоследствии и контролирует, как на уровне транскрипции, так и на уровне трансляции, синтез вителлогенина клетками печени.
Созревание ооцита — это процесс последовательного прохождения двух делений мейоза (делений созревания). При подготовке к первому делению ооцит длительное время находится в фазе диакинеза, когда и происходит его рост и вителлогенез. Начало собственно делений созревания приурочено к достижению самкой половой зрелости и определяется половыми гормонами.
Контроль за процессом созревания яйцеклетки лучше всего изучен у амфибий. У этих животных гонадотропины, находящиеся под контролем гипофиза, действуя на окружающие ооциты фолликулярные клетки, инициируют выделение последними стероидного гормона прогестерона. Подобно другим стероидным гормонам, прогестерон способен диффундировать через плазматические мембраны большинства клеток-мишеней и связываться с внутриклеточными рецепторными белками, регулирующими транскрипцию специфических генов. Однако при созревании ооцита прогестерон, по-видимому, действует иначе. Он связывается с рецепторными белками плазматической мембраны. При этом происходит инактивация плазматической аденилатциклазы, в результате чего снижается концентрация циклического АМФ в цитозоле и соответственно активность цАМФ-зависимой протеинкиназы (А-киназы). Поскольку А-киназа отвечает за фосфорилиро- вание N-концевых участков белков, то ее инактивация приводит к дефосфо- рилированию фактора созревания яйцеклеток (фактор инициации М-фазы, ФИМ), находящегося в цитоплазме. При этом он деблокируется, т. е. переходит в активное состояние.
В норме ФИМ запускает переход из профазы первого деления мейоза в метафазу второго деления. Зрелые ооциты задерживаются на стадии мета- фазы II, когда уровень ФИМ высок. Инактивация А-киназы инициирует активацию небольших количеств ФИМ, которые, в свою очередь, активируют новые порции ФИМ (положительная обратная связь). Замечательное свойство ФИМ — его способность к автокаталитическому саморазмножению, т. е. он может фосфорилировать сам себя и, следовательно, активировать. Фактор созревания вызывает разрушение оболочки ядра ооцита, разрушение ядрышек и миграцию хромосом к будущему анимальному полюсу, где и произойдут деления созревания.
Основная особенность деления созревания в ооцитах состоит в том, что эти деления резко неравномерны. В результате первого деления созревания половина хромосомного набора выталкивается в очень маленькую клетку — редукционное (полярное или направительное) тельце. Впоследствии эта клетка делится на две столь же малые, и никакого участия в дальнейшем развитии они не принимают. Яйцеклетка после выделения первого редукционного тельца называется ооцитом II порядка.
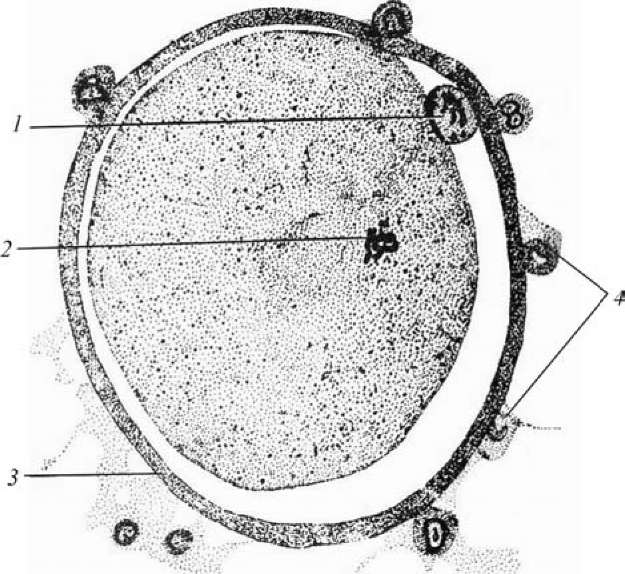 Рис. 8. Ооцит II порядка после овуляции
(по Алмазову, Сутулову, 1978): 1 — первое редукционное тельце; 2 — веретено второго деления созревания; 3 — блестящая зона; 4 — фолликулярные клетки
Рис. 8. Ооцит II порядка после овуляции
(по Алмазову, Сутулову, 1978): 1 — первое редукционное тельце; 2 — веретено второго деления созревания; 3 — блестящая зона; 4 — фолликулярные клетки
|
Второе деление созревания осуществляется путем выделения второго редукционного тельца таких же размеров, как и первое. После его выделения ооцит II порядка превращается в зрелое яйцо (рис. 8).
Количество яиц, созревающих одновременно, редко достигает 15, обычно их бывает меньше, иногда только одно (человек). У большинства животных течение мейоза останавливается на некотором этапе созревания (блок мейоза), а для дальнейшего его протекания требуется оплодотворение яйцеклетки сперматозоидом (исключение составляют морские ежи и некоторые кишечнополостные).
Различают три типа блока мейоза (именно на этом этапе происходит овуляция яйцеклетки):
• на стадии диакинеза (губки, моллюски, отдельные представители плоских, круглых, кольчатых червей, млекопитающие: собака, лиса, лошадь);
• метафазы 1-го деления созревания (губки, немертины, кольчатые черви, насекомые);
• метафазы 2-го деления созревания (хордовые; у летучих мышей блок мейоза наступает в анафазе 2-го деления созревания).
Дата добавления: 2016-12-27; просмотров: 18669;
Поиск по сайту
Узнать еще
- I. Поверхностный аппарат клетки.
- III. ИНДИВИДУАЛЬНО-ПСИХОЛОГИЧЕСКИЕ РАЗЛИЧИЯ БОЙЦОВ
- Бюджетная линия, кривые безразличия.
- ВВЕДЕНИЕ В БОТАНИКУ. ЦИТОЛОГИЯ. КОМПОНЕНТЫ КЛЕТКИ. ЦИТОПЛАЗМА
- Внутренние женские половые органы.
- Внутренние мужские половые органы
- Внутренние различия
- Возрастные и половые особенности черепа

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
