Регуляция транскрипции у прокариот
Свойства генетического кода.

10. Этап собственно трансляции (инициация, элонгация, терминация).
Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК.
После переноса информации с ДНК на матричную РНК начинается синтез белков. Каждая зрелая мРНК несет информацию только об одной полипептидной цепи. Если клетке необходимы другие белки, то необходимо транскрибировать мРНК с иных участков ДНК.
Биосинтез белков или трансляция происходит на рибосомах, внутриклеточных белоксинтезирующих органеллах, и включает 5 ключевых элементов:
· матрица – матричная РНК,
· растущая цепь – полипептид,
· субстрат для синтеза – 20 протеиногенных аминокислот,
· источник энергии – ГТФ,
· рибосомальные белки, рРНК и белковые факторы.
Выделяют три основных стадии трансляции: инициация, элонгация, терминация.
Инициация
Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина.
В начале этой стадии формируются два тройных комплекса:
· первый комплекс – мРНК + малая субъединица + ИФ-3,
· второй комплекс – метионил-тРНК + ИФ-2 + ГТФ.
После формирования тройные комплексы объединяются с большой субъединицей рибосомы. В этом процессе активно участвуют белковые факторы инициации, источником энергии служит ГТФ. После сборки комплекса инициирующаяметионил-тРНК связывается с первым кодоном АУГ матричной РНК и располагается в П-центре (пептидильный центр) большой субъединицы. А-центр (аминоацильный центр) остается свободным, он будет задействован на стадии элонгации для связывания аминоацил-тРНК.
После присоединения большой субъединицы начинается стадия элонгации.
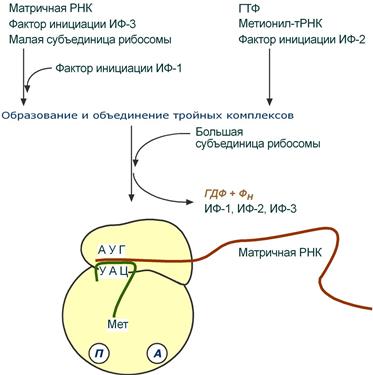
События стадии инициации
Элонгация
Для этой стадии необходимы все 20 аминокислот, тРНК для всех аминокислот, белковые факторы элонгации, ГТФ. Удлинение цепи происходит со скоростью примерно 20 аминокислот в секунду.
Элонгация представляет собой циклический процесс. Первый цикл (и следующие циклы) элонгации включает три шага:
1. Присоединение аминоацил-тРНК (еще второй) к кодону мРНК (еще второму), аминокислота при этом встраивается в А-центр рибосомы. Источником энергии служит ГТФ.
2. Фермент пептидилтрансфераза осуществляет перенос метионина с метионил-тРНК (в П-центре) на вторую аминоацил-тРНК (в А-центре) с образованием пептидной связи между метионином и второй аминокислотой. При этом уже активированная СООН-группа метионина связывается со свободной NH2-группой второй аминокислоты. Здесь источником энергии служит макроэргическая связь между аминокислотой и тРНК.
3. Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы, второй кодон (на рисунке ) становится напротив П-центра, напротив А-центра оказывается третий кодон (на рисунке ). Для этих процессов необходима затрата энергии ГТФ. Так как вместе с мРНК перемещаются закрепленные на ней тРНК, то инициирующая первая тРНК выходит из рибосомы, вторая тРНК с дипептидом помещается в П-центр.
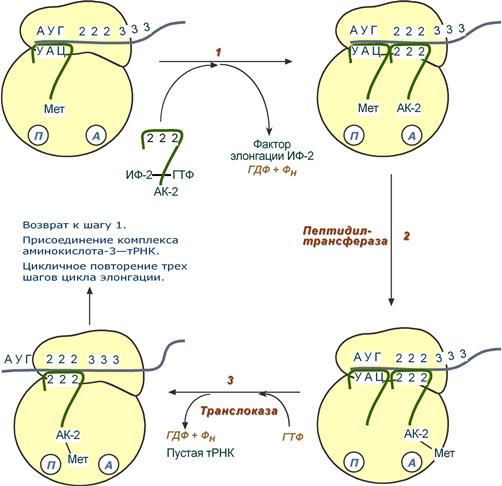
Последовательность событий стадии элонгации
Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А-центр, после чего он смещается в П-центр в транслоказной реакции..
В пустой А-центр входит четвертая аминоацил-тРНК и начинается третий цикл элонгации:
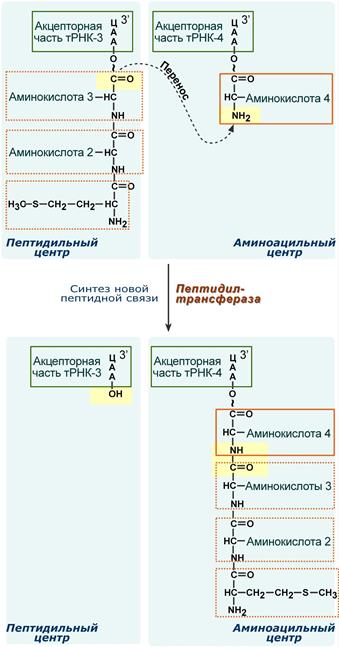
Образование пептидной связи при встраивании четвертой аминокислоты в пептид.
Субъединицы рибосомы, большая часть транспортных РНК и матричная РНК не показаны.
Цикл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь.
Терминация
Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют:
1. Гидролитическое отщепление полипептида от конечной тРНК.
2. Отделение от П-центра последней, уже пустой, тРНК.
3. Диссоциацию рибосомы.
Источником энергии для завершения трансляции является ГТФ.
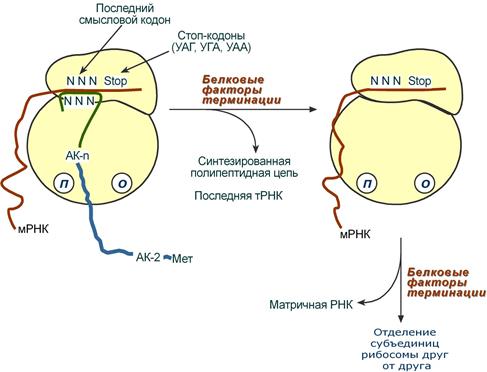
Реакции стадии терминации
Полирибосомы
По причине того, что продолжительность жизни матричной РНК невелика, перед клеткой стоит задача использовать ее максимально эффективно, т.е. получить максимальное количество "белковых копий". Для достижения этой цели на каждой мРНК может располагаться не одна, а несколько рибосом, встающих последовательно друг за другом и синтезирующих пептидные цепи. Такие образования называются полирибосомы.
11. Регуляция биосинтеза белка у эукариот и прокариот на примере лакоперона по теории Жакоба и Моно.
Прокариоты и эукариоты способны осуществлять дифференциальную регуляцию экспрессии генов. Контроль осуществляется за тем, каким генам экспрессироваться, а каким – нет, а также регулируется уровень экспрессии различных генов. Регулироваться может какой-то один или несколько отдельных этапов считывания генетической информации при биосинтезе белка.
Регуляция синтеза белка у прокариот. Общую теорию регуляции синтеза белка разработали французские ученые, лауреаты Нобелевской премии Ф. Жакоб и Ж. Моно. Сущность этой теории сводится к «выключению» и «включению» генов как функциональных единиц (рис. 23).
Согласно теории Ф. Жакоба и Ж. Моно, в биосинтезе белка у бактерий (E. сoli) участвуют, по крайней мере, три типа генов: структурные, ген-регулятор и ген-оператор. Синтез мРНК на структурных генах молекулы ДНК непосредственно контролируется определенным участком, называемым геном-оператором. Ген-оператор локализован на крайнем отрезке структурного гена (структурных генов), регулируемого им. «Считывание» генетического кода, т.е. формирование мРНК, начинается с промотора – участка ДНК, расположенного рядом с геном-оператором и являющегося точкой инициации для синтеза мРНК. Синтезированную молекулу мРНК, кодирующую синтез нескольких разных белков, называют полицистронным (полигенным) транскриптом. Координированный одним оператором одиночный ген или группа генов образует оперон. Деятельность оперона находится под контролирующим влиянием гена-регулятора. Структурные гены и ген-регулятор расположены в разных участках цепи ДНК, поэтому связь между ними осуществляется при помощи белка-репрессора. Репрессор имеет сродство к гену-оператору и обратимо связывается с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка (т.е. функция гена-герулятора состоит в том, чтобы через белок-репрессор прекращать (запрещать) деятельность структурных генов, синтезирующих мРНК). Репрессор, кроме того, обладает способностью строго специфично связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Если такой индуктор соединяется с репрессором, то последний теряет способность связываться с геном-оператором, который, таким образом выходит из под контроля гена-регулятора, и начинается синтез мРНК.
Эукариотические клетки обладают более сложными механизмами регуляции белкового синтеза, так как процессы транскрипции и трансляции разделены не только пространственно ядерной биомембраной, но и во времени. Эта регуляция базируется минимум на шести уровнях сложных биологических процессов, определяющих скорость синтеза и распада генетического продукта (рис. 24).
Для большинства экариотических клеток, как и клеток прокариот, стадия инициации транскрипции является основной. Тем не менее, имеются существенные различия: во-первых, место транскрипции (в ядре) и трансляции (в цитоплазме); во-вторых, активирование транскрипции у эукариот связано с множеством сложных изменений структуры хроматина в транскрибируемой области; в-третьих, в эукариотических клетках положительные регуляторные механизмы превалируют над отрицательными.
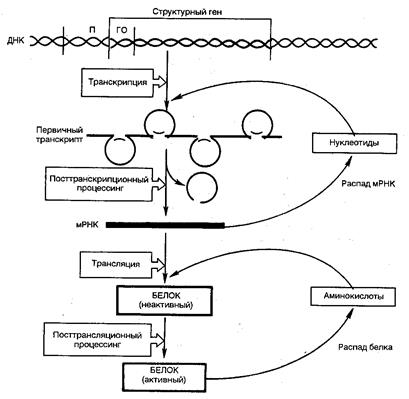
Рис.24. Регуляция экспрессии гена у эукариот
Транскрипция хорошо регулируется
Так как транскрипция связывает ядро – "мозг" клетки, ее "банк знаний" и белки, "рабочих лошадок" клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.
У прокариот и эукариот регуляция транскрипции происходит, естественно, по-разному, хотя некоторые моменты похожи.
Регуляция транскрипции у прокариот
Регуляция биосинтеза белка у прокариот осуществляется на уровне изменения скорости синтеза мРНК. В настоящее время принятатеория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
· оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментовв клетке,
· конституитивные гены – это гены, обеспечивающие основные жизненные функции клетки, "гены домашнего хозяйства". Эти гены работают в клетке всегда, независимо от ее активности и условий,
· индуцибельные гены – это гены, которые активируются при необходимости,
· ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме,
· ген-оператор – участок ДНК, способный связываться с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет.
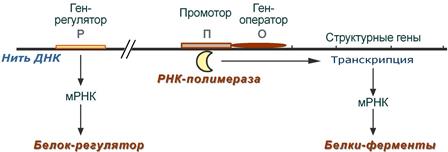
Схема строения и работы оперона
Предложены две схемы регуляции скорости транскрипции: по механизму индукции (лактозный оперон) и по механизму репресии (триптофановый оперон).
Лактозный оперон
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
· активность одного из ферментов катаболизма лактозы низка, если в среде имеется много глюкозы.
· активность этого же фермента резко повышается в обратной ситуации, т.е. при отсутствииглюкозыи при наличии лактозы.
На основании наблюдений была предложена схема регуляции оперона по механизму индукции:
1. При отсутствии лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются.
2. Если глюкозы нет, алактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая ему связаться с геном-оператором и препятствовать работе РНК-полимеразы. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
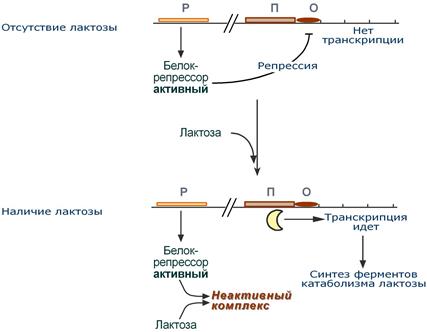
Схема работы лактозного оперона при наличии и отсутствии лактозы
Триптофановый оперон
Триптофановый оперон в целом отвечает за синтез триптофана.
Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии.
1. В отличие от лактозного оперона, белок-репрессор синтезируется в неактивномсостоянии и не может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан.
2. Триптофансоединяется с белком-репрессором и активируетего. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
В этом случае триптофан является репрессором транскрипции.
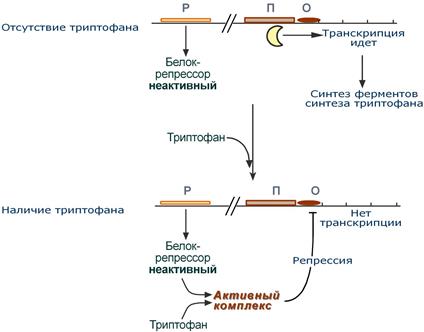
Схема работы триптофанового оперона при наличии и отсутствии триптофана
12. Особенности регуляции биосинтеза белка у эукариот. Регуляторные
области ДНК: энхансеры и сайленсеры.
Дата добавления: 2022-05-27; просмотров: 1021;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
