Специальные межклеточные соединения (контакты)
Электронно-микроскопические исследования позволили обнаружить у живых организмов целый ряд специальных межклеточных структур - контактов, или соединений, которые служат для прикрепления клеток к друг другу (плотные контакты ) и (или) выполняют коммуникационные функции ( щелевые соединения, десмосомы, плазмодесмы).
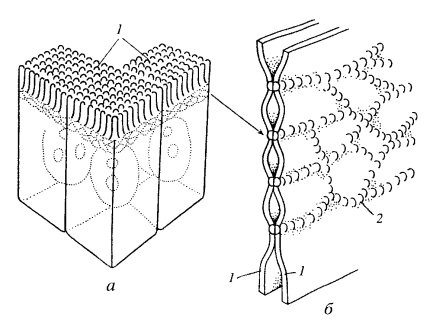
Запирающий (плотный) контакт,характерен для однослойных эпителиальных тканей животных . В зоне контакта плазматические мембраны клеток максимально сближены. Часто видна трехслойность мембраны в этом контакте: два внешних осмофильных слоя обеих мембран как бы сливаются в один общий слой толщиной 2-3 нм. Слияние мембран происходит не по всей площади в зоне плотного контакта, а только в определенных участках (точечно) (рис. 2).
|
| Рис. 2. Схематичное изображение плотного контакта клеток кишечного эпителия а — расположение плотного соединения (вставочная пластинка) на клетках (1) кишечного эпителия; б — трехмерная схема участка плотного соединения: 1 — плазматические мембраны соседних клеток, 2 — глобулы белка окклюдина |
В зоне плотного контакта участки соприкосновения мембран образованы специфическими интегральными глобулярными белками - окклудином и клаудином. На клеточной поверхности они выглядят в виде поясков, которые пересекаются и образуют в ткани сеть. Такие структуры характерны, например, для эпителиев, особенно железистых и кишечных. В последнем случае плотный контакт образует сплошную зону слияния плазматических мембран, опоясывающую клетку в апикальной ( смотрящей в просвет кишечника) ее части. Таким образом, каждая клетка пласта как бы обведена лентой этого контакта. Такие структуры получили у морфологов название замыкающих пластинок. В эпителиальной ткани плотные контакты не только механически соединяют клетки друг с другом, но и она запирают, перегораживают межклеточные полости, изолируя их (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Зона плотного контакта практически не проницаема не только для макромолекул, но и для малых молекул и ионов. Описанные контакты встречаются между всеми типами однослойного эпителия (эндотелий, мезотелий, эпендима).
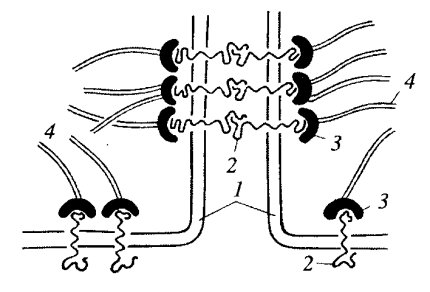
Заякоривающие,или сцепляющие, соединения,или контакты, так называются потому, что они соединяют не только плазматические мембраны соседних клеток, но и связываются с фибриллярными элементами цитоскелета (рис. 3). Для этого рода соединений характерным является наличие двух типов белков. Первый тип представлен трансмембранными линкерными (связующими) белками, которые участвуют или в собственно межклеточном соединении или в соединении плазмалеммы с компонентами внеклеточного матрикса (базальная мембрана эпителиев, внеклеточные структурные белки соединительной ткани).
|
| Рис. 3. Схематичное изображение строения заякоривающего адгезивного контакта 1 — плазматическая мембрана; 2 — трансмембранные линкерные гликопротеиды; 3 — внутриклеточные белки сцепления; 4 — элементы цитоскелета |
Ко второму типу относятся внутриклеточные белки, соединяющие, или заякоривающие, мембранные элементы такого контакта с цитоплазматическими фибриллами цитоскелета. К заякоривающим соединениям относятся межклеточные сцепляющие точечные контакты, сцепляющие ленты, фокальные контакты, или бляшки сцепления; все эти контакты связываются внутри клеток с актиновыми микрофиламентами.
Другую группу заякоривающих межклеточных соединений составляют десмосомыи полудесмосомы; они связываются с другими элементами цитоскелета — с промежуточными филаментами.
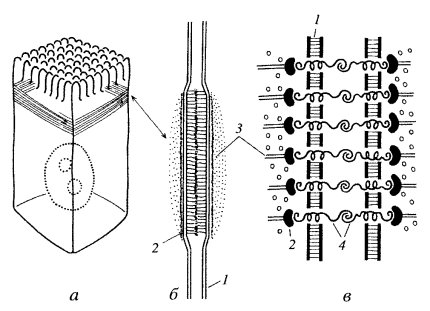
Межклеточные точечные сцепляющие соединения обнаружены у многих неэпителиальных тканей, но более отчетливо описана структура сцепляющих (адгезивных) лентв однослойных эпителиях (рис. 3). Эта структура опоясывает весь периметр эпителиальной клетки, подобно тому как это происходит в случае плотного соединения. Чаще всего такой поясок, или лента, лежит ниже плотного соединения. В этом месте плазматические мембраны не сближены, а даже несколько раздвинуты на расстояние 25—30 нм, и между ними видна зона повышенной плотности. Это не что иное, как места взаимодействия трансмембранных гликопротеидов, которые специфически сцепляются друг с другом и обеспечивают механическое соединение мембран двух соседних клеток. Эти линкерные белки относятся к Е-кадгеринам — белкам, обеспечивающим специфическое узнавание клетками однородных мембран. Разрушение этого слоя гликопротеидов приводит к обособлению отдельных клеток и к разрушению эпителиального пласта. С цитоплазматической стороны около мембраны видно скопление какого-то плотного вещества, к которому примыкает слой тонких (6—7 нм) филаментов, лежащих вдоль плазматической мембраны в виде пучка, идущего по всему периметру клетки. Тонкие филаменты относятся к актиновым фибриллам, они связываются с плазматической мембраной посредством белков катенина, винкулина и α-актинина, образующих плотный околомембранный слой.
|
| Рис. 4. Адгезивный (сцепляющий) поясок (лента) а — расположение пояска в клетке; 6 — вид на поперечном срезе; 1 — плазматическая мембрана; 2 — слой винкулина; 3 — актиновые микрофиламенты; 4 — линкерные гликопротеиды |
Функциональное значение такого ленточного соединения заключается не только в механическом сцеплении клеток друг с другом: при сокращении актиновых филаментов в ленте может изменяться форма клетки. Кооперативное сокращение актиновых фибрилл во всех клетках эпителиального пласта может вызвать изменение его геометрии, например сворачивание в трубку, подобно тому, что происходит при образовании нервной трубки у эмбрионов позвоночных.
Фокальные контакты,или бляшки сцепления,встречаются у многих клеток и особенно хорошо изучены у фибробластов. Они построены по общему плану со сцепляющими лентами, но выражены в виде не больших участков — бляшек — на плазмалемме. В этом случае трансмембранные линкерные белки-интегрины специфически связываются с белками внеклеточного матрикса (например, с фибронектином). Со стороны цитоплазмы эти же гликопротеиды связаны с примембранными белками, куда входит и винкулин, который в свою очередь связан с пучком актиновых филаментов. Функциональное значение фокальных контактов заключается как в закреплении клетки на внеклеточных структурах, так и в создании механизма, позволяющего клеткам перемещаться.
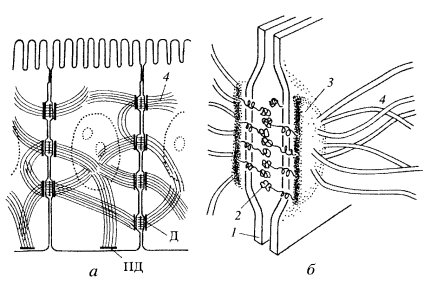
Десмосомы — структуры в виде бляшек или кнопок, также соединяют клетки друг с другом (рис. 5). В межклеточном пространстве здесь также виден плотный слой, представленный взаимодействующими интегральными мембранными кадгеринами — десмоглеинами, которые сцепляют клетки друг с другом. С цитоплазматической стороны к плазмалемме прилежит слой белка-десмоплакина, с которым связаны промежуточные филаменты цитоскелета. Десмосомы встречаются чаще всего в эпителиях, в этом случае промежуточные филаменты содержат кератины. Клетки сердечной мышцы — кардиомиоциты, содержат десминовые фибриллы в составе десмосом. В эндотелии сосудов в состав десмосом входят виментиновые промежуточные филаменты.
|
| Рис. 5. Расположение десмосом на мембранах контактирующих клеток 1 — плазматическая мембрана; 2 — десмоглеиновый слой; 3 — слой десмоплакина; 4 — промежуточные филаменты. Д — десмосома; ПД — полудесмосома |
Полудесмосомы в принципе сходны по строению с десмосомой, но представляют собой соединение клеток с межклеточными структурами. Так, в эпителиях линкерные гликопротеиды (интегрины) десмосомы взаимодействуют с белками так называемой базальной мембраны, куда входят коллаген, ламинин, протеогликаны и др.
Функциональная роль десмосом и полудесмосом сугубо механическая — они сцепляют клетки друг с другом и с подлежащим внеклеточным матриксом прочно, что позволяет эпителиальным пластам выдерживать большие механические нагрузки. Подобно этому десмосомы прочно связывают друг с другом клетки сердечной мышцы, что позволяет им выполнять огромную механическую нагрузку, оставаясь связанными в единую сокращающуюся структуру.
В отличие от плотного контакта все типы сцепляющих контактов проницаемы для водных растворов и не играют никакой роли в ограничении диффузии.
Щелевые контакты участвуют в прямой передаче химических веществ из клетки в клетку и соответственно, выполняют важную роль при функционировании клеток специализированных тканей, при развитии новых тканей организма, при дифференцировке клеток. Характерным для этого типа контактов является сближение плазматических мембран двух соседних клеток на расстояние 2—3 нм. Дальнейший прогресс в расшифровке его строения был достигнут при использовании метода замораживания-скалывания. Было покаазано, что зоны щелевых контактов (размером от 0,5 до 5 мкм) усеяны гексагонально расположенными (с периодом 8—10 нм) частицами 7—8 нм в диаметре, имеющими в центре канал около 2 нм шириной. Эти частицы получили название коннексонов(рис. 6). В зонах щелевого контакта может быть от 10 до нескольких тысяч коннексонов в зависимости от функциональных особенностей клеток. Коннексоны были выделены препаративно, оказалось, что они состоят из шести субъединиц коннексина— белка с молекулярной массой около 30 кД. Коннексины двух мембран образуют цилиндрический агрегат — коннексон, который пронизывает мембраны кон тактирующих клеток. Коннексоны играют роль прямых межклеточных каналов, по которым ионы и низкомолекулярные вещества могут диффундировать из клетки в клетку.
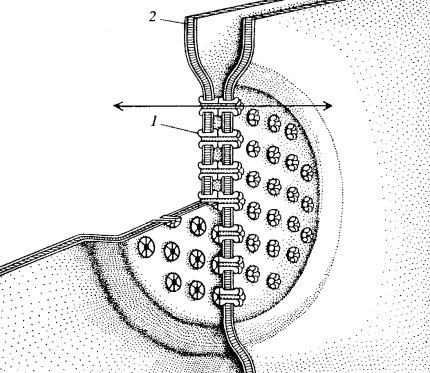
|
| Рис. 6. Схема щелевого соединения между двумя клетками 1 — коннексон; 2 — плазматическая мембрана. Стрелкой обозначен белковый канал, образованный коннексинами |
Через щелевые контакты могут транспортироваться вещества с молекулярной массой не более 1500 Да и диаметром не более 1,5 нм (ионы, аминокислоты, нуклеотиды, сахара, витамины, стероиды, гормоны). Такая способность щелевых контактов служить местом транспорта низкомолекулярных соединений используется в тех клеточных системах, где нужна быстрая передача электрического импульса (волны возбуждения) от клетки к клетке без участия нервного медиатора. Так, все мышечные клетки миокарда сердца связаны с помощью щелевых контактов. Это создает условие для синхронного сокращения огромного количества клеток. При росте культуры эмбриональных сердечных мышечных клеток (миокардиоцитов) некоторые клетки в пласте начинают независимо друг от друга спонтанно сокращаться с разной частотой, и лишь после образования между ними щелевых контактов они начинают биться синхронно, как единый сокращающийся пласт клеток. Таким же способом обеспечивается совместное сокращение гладкомышечных клеток в стенке матки.
Щелевые контакты могут служить целям метаболической кооперации между клетками, обмениваясь различными молекулами, гормонами, цАМФ или метаболитами. У ранних эмбрионов позвоночных, начиная с восьмиклеточной стадии, большинство клеток связано друг с другом щелевыми контактами. По мере дифференцировки эмбриона щелевые контакты между всеми клетками исчезают и остаются только между группами специализирующихся клеток. Например, при образовании нервной трубки связь клеток этой структуры с остальным эпидермисом прерывается и они разобщаются.
Плазмодесмы. Этот тип межклеточных контактов характерен для растений. Плазмодесмы представляют собой тонкие трубчатые цитоплазматические каналы, соединяющие две соседние клетки. Диаметр этих каналов обычно составляет 20—40 нм. Ограничивающая эти каналы мембрана непосредственно переходит в плазматические мембраны соседствующих клеток. Плазмодесмы проходят сквозь клеточную стенку, разделяющую клетки (рис. 7). Таким образом, у некоторых растительных клеток плазмодесмы соединяют гиалоплазму соседних клеток, поэтому формально здесь нет полного разграничения, отделения тела одной клетки от другой, это скорее представляет собой синцитий: объединение многих клеточных территорий с помощью цитоплазматических мостиков. Внутрь плазмодесм могут проникать мембранные трубчатые элементы, соединяющие цистерны эндоплазматического ретикулума соседних клеток. Образуются плазмодесмы во время деления клетки, когда строится первичная клеточная оболочка. У только что разделившихся клеток количество плазмодесм может быть большим (до 1000 на клетку), при старении клеток их количество снижается по мере увеличения толщины клеточной стенки.
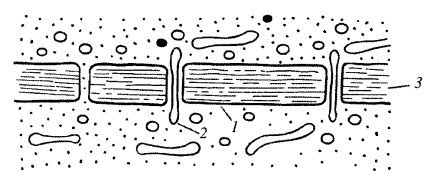
|
| Рис. 7. Схема строения плазмодесмы на между двумя растительными клетками 1 — плазматическая мембрана; 2 — мембрана десмотубулы; 3 — клеточная стенка |
Функция плазмодесм заключается в обеспечении обеспечивается межклеточной циркуляция растворов, содержащих небольшие молекулы (до 800 Да), ионы . В частности, по плазмодесмам могут перемещаться небольшие липидные капли. Заражение растительных клеток вирусами также может происходить через плазмодесмы.
Вопросы для самоконтроля
1. Дайте определение термину «информация». Перечислите формы и способы передачи информации в живых системах.
2. Сформулируйте и объясните закон Стивенса; закон Вебера-Фехнера?
3. В каких единицах измеряется количество информация ? Дайте объяснения.
4. Каков абсолютный порог чувствительности для уха, для глаза человека?
5. Какова пропускная способность информационных каналов в организме человека: уха, глаза?
6. Перечислите и охарактеризуйте виды межклеточных контактов у живых организмов.
7. Что означает термин «коннексон» ?
Лекция 10.
Дата добавления: 2018-05-10; просмотров: 3392;
Поиск по сайту
Узнать еще
- I. Реакции присоединения
- VI. ЩЕЛЕВЫЕ И СПЕЦИАЛЬНЫЕ ТИПЫ АНТЕНН
- Автомобильной службы части (соединения) после получения задачи
- Автомобильные топлива, смазочные материалы и специальные жидкости.
- Автотехническим обеспечением части (соединения)
- АЛИЦИКЛИЧЕСКИЕ СОЕДИНЕНИЯ
- Алюминий и его соединения: получение, физические и химические свойства. Медико-биологическое значение алюминия.
- анатомия костей скелета. соединениякостей

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
